Пропоную вашій увазі урок "Ознака як результат взаємодії генів. Поняття про зчеплення генів і кросинговер". Зчеплене успадкування полягає в тому, що гени, які локалізовані в одній хромосомі спадкуються переважно разом або зчеплено. Зчеплення між генами може бути повне, якщо між ними не відбувається кросинговер. При повному зчепленні дигетерозиготи формують лише два типи некросовесних гамет у співвідношенні 1:1. При наявності кросинговера дигетерозиготи формують чотири типи гамет, з яких два типи некросоверні і два типи кросоверні. Частка кросоверних гамет незначна. Вона дорівнює відстані між зчепленими генами. Відстань між генами позначається морганидами або у відсотках кросинговера.
Конструктор уроків
- Всеосвіта›
- Бібліотека уроків›
- Біологія›
- Ознака як результат взаємодії генів. Поняття про зчеплення генів і кросинговер

Урок:
Ознака як результат взаємодії генів. Поняття про зчеплення генів і кросинговер
31.01.2022
Опис уроку (учням цей опис не показується):
Вміст уроку:
1
2
3
4
Опис, який учні побачать перед початком уроку
Бажаю успіху!
1
1 з 12 балів
Учені, намагаючись повторювати досліди Г. Менделя на різних організмах (не тільки на рослинах, а й на тваринах), часто спостерігали варіанти розщеплення, які не відповідали очікуваним згідно зі встановленими ним законами спадковості. Деякі з причин цього ми згадували раніше: це проміжний характер успадкування, кодомінантність, вплив летальних алелів. Крім того, однією з основних причин відхилень варіантів розщеплення від передбачених законами спадковості є явище зчепленого успадкування.
Причини прояву явища зчепленого успадкування
Закон незалежного комбінування ознак базується на таких положеннях:
розвиток різних варіантів ознак зумовлений алельними генами, які розташовані в однакових локусах у гомологічних хромосомах;
гамети та інші гаплоїдні клітини, які мають по одній хромосомі з кожної пари гомологічних хромосом, несуть лише один алельний ген з певної їхньої кількості;
гени, які контролюють розвиток ознак, що успадковуються незалежно, розташовані в негомологічних хромосомах.
Гени однієї хромосоми утворюють окрему групу зчеплення.
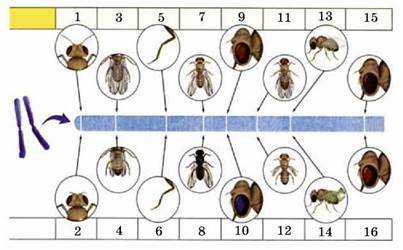
Група зчеплення, позначена на генетичній карті хромосоми дрозофіли (цифрами позначені відповідні алельні гени): 1 - довгі вусики; 2 - короткі вусики; 3 - довгі крила; 4 - короткі крила; 5 - довгі лапки; 6 - короткі лапки; 7 - сіре тіло; 8 - чорне тіло; 9 - червоні очі; 10 - фіолетові очі; 11 - нормальні крила; 12 - редуковані крила; 13 - рівні крила; 14 - вигнуті крила; 15 - червоні очі; 16 - коричневі очі
Генетичними картами хромосом називають схеми, на яких показано порядок розташування генів у хромосомі та відносні відстані між ними. Такі карти мають не тільки теоретичний інтерес, а й широке використання у селекції та генній інженерії.
Кількість спадкових ознак організмів значно перевищує кількість їхніх хромосом у гаплоїдному наборі. Наприклад, у гаплоїдному наборі мухи дрозофіли усього чотири хромосоми, а кількість структурних генів, які визначають різні спадкові ознаки, становить близько 10 тис. Відповідно, у кожній хромосомі є не один, а багато генів. Таким чином, одночасно з ознаками, які успадковуються незалежно одна від одної, існують і такі, які успадковуються зчеплено, оскільки контролюються генами, розташованими в одній хромосомі.
На малюнку вище наведено частину генетичної карти хромосоми мухи дрозофіли з позначенням груп зчеплення. Кількість таких груп в організмів певного виду дорівнює кількості нестатевих хромосом (аутосом) і типів статевих хромосом. Наприклад, у самки дрозофіли кількість груп зчеплення становить 4, тоді як у самців - 5 (оскільки X- та Y-хромосоми відрізняються за набором генів). Так само у жінок кількість груп зчеплення становить 23, а у чоловіків - 24.
Явище зчепленого успадкування досліджував видатний американський генетик Т.Х. Морган. Він вдало обрав об’єкт для експериментів - муху дрозофілу. Цих комах легко утримувати в лабораторіях, для них характерні висока плодючість і швидка зміна поколінь (за оптимальних умов утримання нове покоління з’являється кожні півтора-два тижні), невелика кількість хромосом (у диплоїдному наборі - 8). Оскільки дрозофіла розвивається з повним перетворенням, є можливість вивчати активність генів на різних фазах розвитку.
Явище зчепленого успадкування Т.Х. Морган довів за допомогою такого досліду. Самців дрозофіли, гомозиготних за домінантними алелями забарвлення тіла (сіре) і форми крил (нормальна), він схрестив із самками, гомозиготними за рецесивними алелями відповідних генів (чорне забарвлення тіла й недорозвинені крила). Усі гібриди першого покоління, отримані від такого схрещування, були гетерозиготними за обома генами, мали сіре забарвлення тіла і крила нормальної форми. Щоб з’ясувати генотип гібридів першого покоління, Т.Х. Морган застосував аналізуюче схрещування: схрестив їх з особинами, гомозиготними за рецесивними алелями відповідних генів.
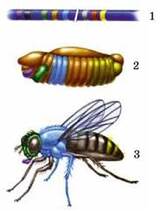
Дрозофіла - класичний об’єкт генетичних досліджень: 1 - ділянка хромосоми; 2 - лялечка; 3 - доросла комаха (кольорами схематично показано гени й ознаки, розвиток яких вони визначають)
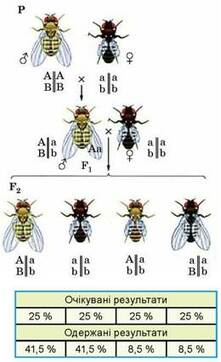
Зчеплене успадкування деяких ознак (забарвлення тіла та форми крил) у дрозофіли
Теоретично серед нащадків, отриманих від такого схрещування, можна було очікувати два варіанти розщеплення. Якби гени, що визначають забарвлення тіла і форму крил, були розташовані в негомологічних хромосомах розщеплення мало бути таким: 25 % особин із сірим тілом і нормальною формою крил, 25 % - із сірим тілом і недорозвиненими крилами, 25 % - із чорним тілом і нормальними крилами і 25 % - із чорним тілом і недорозвиненими крилами (тобто в співвідношенні 1 : 1 : 1 : 1). Але якби ці гени були розташовані в одній хромосомі й успадковувалися зчеплено, то серед нащадків можна було б очікувати 50 % особин із сірим тілом і нормальною формою крил і 50 % - із чорним тілом і недорозвиненими крилами (тобто у співвідношенні 1 : 1).
Практично отримані результати були такими: 41,5 % нащадків мали сіре тіло і нормальну форму крил, 41,5 % - чорне тіло і недорозвинені крила, 8,5 % - сіре тіло і недорозвинені крила і 8,5 % - чорне тіло і нормальну форму крил. Отримане внаслідок аналізуючого схрещування розщеплення за фенотипами, подібними до батьківських форм, наближувалося 1 : 1 (як у разі зчепленого успадкування). Але серед нащадків були й два інші варіанти фенотипів, які можна було б очікувати тоді, коли б гени забарвлення тіла і форми крил розташовувалися в негомологічних хромосомах та успадковувалися незалежно. Проте їхня кількість була занадто малою.
Як Т.Х. Морган припустив, що гени, які визначають забарвлення тіла мух і форму їхніх крил, розташовані в одній хромосомі, тож успадковуються зчеплено. Але під час утворення статевих клітин, у процесі мейозу, гомологічні хромосоми здатні обмінюватися своїми гомологічними ділянками (явище кросинговеру).
Дослідження кросинговеру дали можливість виявити закономірності, які стали підґрунтям для хромосомної теорії спадковості:
гени розташовані в хромосомі по її довжині в лінійному порядку, тобто один за одним;
різні хромосоми мають неоднакові набори генів, тобто кожна з негомологічних хромосом має свій унікальний набір генів;
кожний ген розташований у хромосомі у певному місці - локусі; алельні гени займають у гомологічних хромосомах однакові локуси;
усі гени однієї хромосоми утворюють групу зчеплення, завдяки чому відбувається зчеплене успадкування деяких ознак;
сила зчеплення між двома генами, розташованими в одній хромосомі, обернено пропорційна відстані між ними; що більша ця відстань, то частіше відбувається кросинговер;
у довгих хромосомах є гени, розташовані на їхніх різних полюсах; ознаки, які вони визначають, можуть успадковуватися незалежно, так само як і ознаки, що визначаються генами, розташованими в негомологічних хромосомах;
частота кросинговеру між двома генами, розташованими в одній хромосомі, є величиною відносно постійною для кожної конкретної пари генів; утім, на неї можуть впливати деякі фактори зовнішнього і внутрішнього середовища; у деяких організмів виявлено залежність частоти кросинговеру від віку або статі;
відносну постійність частоти кросинговеру між різними парами генів однієї групи зчеплення використовують як показник відстані між окремими генами, а також для визначення послідовності їх розташування в хромосомі. Зокрема, визначення частот кросинговеру між різними парами генів використовують для складання генетичних карт хромосом;
зчеплення між генами однієї групи порушується внаслідок обміну ділянками гомологічних хромосом у профазі першого мейотичного поділу (процес кросинговеру).
Відстань між двома генами однієї хромосоми вимірюють у сантиморганах (скорочено - сМ) (цю одиницю виміру названо на честь Т.Х. Моргана): 1 сантиморган відповідає такій відстані між двома генами, за якої процес кросинговеру відбувається в 1 гаметі з кожної сотні, тобто з імовірністю 1 % (1 сМ = 1 %). Слід зазначити, що за відстані в 50 морганід і більше гени успадковуються незалежно, так, наче вони розташовані в негомологічних хромосомах.
2
3 з 12 балів
Дайте відповіді на запитання:
Із чим пов’язане явище зчепленого успадкування ознак?
Чим визначається кількість груп зчеплення в організмів певного виду?
Яке біологічне значення кросинговеру?
.png)
3
6 з 12 балів

31 січня 2022
 Чернишова Світлана Володимирівна
Чернишова Світлана Володимирівна
Чернишова Світлана Володимирівна
Чернишова Світлана Володимирівна
Пропоную вашій увазі тест "Явище зщепленого успадкування". Часто спостерігаються варіанти розщепленн...
Біологія
9 клас
4
2 з 12 балів
Поміркуйте
Дрозофіла стала класичним об’єктом для генетичних досліджень. Поміркуйте, які ще тварини могли б стати класичним об’єктом генетичних досліджень.
Опис, який учні побачать після проходження уроку
Молодець!
Рефлексія від 4 учнів
Сподобався:
0
Так: 3
Ні: 1
Зрозумілий:
0
Так: 3
Ні: 1
Потрібні роз'яснення:
0
Ні: 4
Так: 0