Конструктор уроків
- Всеосвіта›
- Бібліотека уроків›
- Біологія›
- Конспект на контрольну роботу

Урок:
Конспект на контрольну роботу

25.09.2023
Вміст уроку:
1
2
3
4
Опис, який учні побачать перед початком уроку
Конспект параграфів 7 - 17
1
КОНСПЕКТ параграфів 7-17
ПАРАГРАФ 7 НЕКЛІТИННІ ФОРМИ ЖИТТЯ
Неклітинні форми життя — це автономні структури на молекулярному рівні організації життя, до яких відносять віроїди, пріони та віруси. Спільними ознаками цих форм життя є: а) субмікроскопічні розміри; б) відсутність клітинної будови; в) внутрішньоклітинний паразитизм; в) здатність зберігати свої ознаки та змінюватися під упливом умов; г) здатність до самовідтворення.
Віроїди — неклітинні форми життя у вигляді одноланцюгової кільцеподібної РНК, що не кодує білків. Віроїди було відкрито в 1971 р. американським біологом Т. О. Дінером. Ці субмікроскопічні частинки захисної білкової оболонки не мають, їхня РНК містить 250—375 нуклеотидів і має масу від 86—130 кДа. Віроїди потрапляють у клітини організму хазяїна під час його вегетативного розмноження, за допомогою комах або механічним шляхом у разі пошкодження. Віроїди можуть завдавати значних збитків рослинництву, оскільки спричиняють віроїдні захворювання рослин (веретеноподібність бульб картоплі, карликовість хризантем, каданг-каданг кокосових пальм, хвороба жовтих плям рису та ін.).
Пріони — неклітинні форми життя, що є білковими частинками без нуклеїнової кислоти. Пріони складаються із особливого білка, який існує у вигляді двох форм: нормальний білок масою близько 25 400 а. о. м. та аномальний, що відрізняється вторинною структурою і здатний перетворювати нормальний білок на собі подібний (іл. 10). Особливостями пріонів є відсутність власного геному, висока стійкість до температури, ультрафіолету, радіації. Пріони не розпізнаються імунною системою як чужорідні білки й не провокують імунної відповіді. Відкрив пріони в 1982 р. американський біохімік С. Прузінер. На сьогодні пріони знайдено в клітинах бактерій, дріжджів й ссавців. Описано понад 10 небезпечних пріонних хвороб тварин й людини (коров’ячий сказ, скрейпі овець, губчаста енцефалопатія котячих, фатальне сімейне безсоння, хвороба куру та ін.).
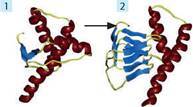
Іл. 10. Форма пріонів: 1 — нормальна форма білка; 2 — аномальна форма білка
ВІРУСИ (лат. Virus — отрута) — це неклітинні форми життя, які є внутрішньоклітинними абсолютними паразитами (іл. 11). Наразі близько 5 000 видів вірусів ідентифіковано, тоді як вчені вважають, що існує понад мільйон видів вірусів. Відкрито віруси в 1892 р. російським ботаніком Д. Івановським (1864—1920). Розміри більшості вірусів — у межах від 20 до 300 нм, хоча є серед них й справжні велетні серед неклітинних форм життя. Це мега- й мімівіруси з розмірами близько 600 нм.
Іл. 11. Вірусні частинки: 1 — герпесу; 2 — грипу; 3 — тютюнової мозаїки; 4 — бактеріофаг
Віруси мають капсид з білкових частинок — капсомерів і нуклеїнову кислоту — серцевину. Структуру, що містить нуклеїнову кислоту і капсид, називають нуклеокапсидом. Повністю сформована інфекційна вірусна частинка називається віріоном. Унікальна властивість вірусів — різноманітність організації генетичного матеріалу. Вірусна нуклеїнова кислота може бути представлена одно- або дволанцюговими молекулами ДНК (ДНК-вмісні віруси) чи РНК (РНК-вмісні віруси). Для вірусів характерна специфічність, що полягає в можливості взаємодії вірусів лише з певними рецепторами клітинних мембран.
Віруси є внутрішньоклітинними паразитами архей, бактерій й еукаріотів і поза клітинами жодних ознак життя не виявляють. Для свого відтворення віруси використовують ресурси клітини-хазяїна, чим спричиняють вірусні захворювання. Основними систематичними одиницями для вірусів є порядок, родина, рід та вид. Категорії царства та відділів, бінарна номенклатура для вірусів не застосовуються.
Отже, неклітинні форми життя є складовою частиною біосфери, оскільки побудовані з органічних речовин, мають біотичні взаємовідносини, їм притаманні спадковість, мінливість, вони здатні до самовідтворення та ін.
Як віруси взаємодіють з клітинами?
Віруси не можуть самостійно поширюватись у природі, й допомагають їм у цьому насамперед живі організми: комахи (наприклад, попелиці переносять близько 160 різних арбовірусів), кліщі, ґрунтові нематоди, рослини-паразити (наприклад, повитиця переносить близько 50 вірусів рослин), людина (для вірусів грипу).
Шляхи проникнення вірусів до організму хазяїна бувають різними. Це: 1) повітряно-краплинний шлях (віруси грипу, вітряної віспи, кору); 2) з їжею та водою (вірус ящуру з молоком ураженої корови, гепатит А); 3) крізь покриви тіла (віруси герпесу, сказу, папіломи); 4) під час переливання крові та хірургічних операцій (вірус гепатиту В); 5) статевим шляхом (вірус герпесу, СНІДу); 6) за участі переносників (кліщів — вірус тайгового енцефаліту, комарів — вірус лихоманки Зіка); 7) з пилком рослин (віруси мозаїки картоплі).
Залежно від властивостей вірусу і клітини-хазяїна, а також умов середовища розрізняють такі основні типи взаємодії вірусів із клітиною: а) продуктивна інфекція — розмноження вірусу призводить до руйнування і загибелі клітини; б) абортивна інфекція — взаємодія, за якої клітини залишаються живими, а дозрілі віріони не утворюються; в) вірогенія — геном вірусу поєднується з генетичним апаратом клітини і під час поділу клітини може тривалий час передаватися дочірнім клітинам; через деякий час за певних умов починає розмножуватися і спричиняє загибель клітини.
Отже, взаємодії вірусів із клітинами мають різноманітний та складний характер.
ПАРАГРАФ 8 РОЛЬ ВІРУСІВ У ПРИРОДІ Й ЖИТТІ ЛЮДИНИ
У життєвому циклі вірусів чергуються дві фази — позаклітинна (віріон) і внутрішньоклітинна (іл. 13). Фаза віріона характеризується відсутністю будь-яких проявів життя, а упродовж внутрішньоклітинної фази життєвого циклу виявляються властивості вірусів, що закодовані в їхньому геномі, й відбувається їхнє відтворення. Незалежно від типу генетичного матеріалу віруси мають подібні загальні стадії життєвого циклу.
![]()
Іл. 13. Життєвий цикл фагів: 1 — адсорбція бактеріофага; 2 — впорскування ДНК у клітину; 3 — синтез вірусних ДНК і білків; 4 — збирання вірусних частинок; 5 — вихід із клітини
Адсорбція вірусів — це прикріплення віріонів до поверхневого апарату клітини-хазяїна. Між білками вірусів й рецепторами клітини виникають електростатичні взаємодії, що забезпечують тимчасове нестійке приєднання віріонів.
Проникнення в клітину в різних вірусів здійснюється різними шляхами: а) ендоцитозу (віропексису) з утворенням внутрішньоклітинних вакуолей; б) злиттям плазматичної мембрани з оболонкою вірусу; в) шляхом впорскування (наприклад, у бактеріофагів) тощо.
«Роздягання» вірусів у клітині забезпечує звільнення нуклеїнової кислоти вірусу від захисної білкової оболонки для наступної реалізації вірусного геному. У фагів цієї стадії немає, оскільки білкова оболонка залишається на поверхні бактеріальної клітини.
Відтворення компонентів вірусних частинок забезпечують три процеси: транскрипція, трансляція й реплікація. Транскрипція полягає в переписуванні інформації з вірусних ДНК чи РНК та утворенні іРНК, що відбувається в ядрі (нуклеоїді). Трансляцією називається процес переведення інформації іРНК у послідовність амінокислот молекул білків. Відбувається трансляція на рибосомах за допомогою тРНК й ферментів. Реплікація — синтез молекул нуклеїнової кислоти, що каталізується переважно клітинними ферментами. Таким чином, відтворення вірусів відбувається відокремлено, неодночасно і в різних частинах клітини. Такий спосіб розмноження вірусів називають роз’єднаним (диз’юнктивним).
Збирання віріонів відбувається усередині клітин або шляхом відбруньковування в зовнішнє середовище. Вихід вірусів із клітин може супроводжуватися збереженням життєдіяльності клітини або її руйнуванням і загибеллю.
Отже, найзагальнішими особливостями життєвого циклу вірусів є наявність двох фаз, використання ресурсів клітини та роз’єднаний спосіб розмноження.
Яка роль вірусів в еволюції організмів?
Все живе на Землі насичено різними вірусами, що свідчить про їхні необхідність у живій природі та важливі функції у біосфері. Віруси відіграють ключову біологічну роль на всіх рівнях організації життя. Вони визначають еволюцію організмів завдяки своїй патогенності, здатності до антагоністичних і симбіотичних відносин та участі в горизонтальному перенесенні генів.
Патогенні віруси є рушійною силою еволюції імунної системи. Кожен живий організм, від кишкової палички, хлорели, дріжджів до синього кита чи секвої, може інфікуватися принаймні одним, а зазвичай кількома вірусами. Вірусними захворюваннями рослин є жовтяниця, мозаїчні хвороби, строкатолистість, готика та ін. (іл. 14). У людини віруси спричиняють гепатит В, простий герпес, папілому, натуральну віспу (ДНК-вмісні вірусні інфекції), грип, гепатит А, енцефаліт, кір, свинку, поліомієліт, сказ, СНІД (РНК-вмісні вірусні інфекції) та ін. Найпоширенішими вірусними захворюваннями тварин є ящур, чумка собак, чумка курей та ін.

Іл. 14. Жовтяниця буряка (1) і мозаїка томатів (2)
Вірусна інфекція є потужною силою, котра формує механізми захисту хазяїв. Наприклад, модифікації поверхні клітин з метою запобігання проникненню бактеріофага, система CRISPR бактерій та архей, гуморальний й клітинний імунітет людини тощо. У процесі еволюції віруси виробляють свої способи захисту від імунної системи. Наприклад, вони здатні блокувати роботу NK-кілерів, інтерферону, Т-лімфоцитів.
Завдяки патогенності віруси регулюють чисельність своїх хазяїв (наприклад, ДНК-віруси морських водойм регулюють кількість бактерій) і впливають на процес фотосинтезу в Світовому океані, кругообіг Карбону і навіть на погоду. Так, біогенні сульфуровмісні гази вивільняються через те, що віруси вбивають певні види водоростей. Потім в атмосфері внаслідок окиснення утворюються кислотні краплинки, що слугують ядрами конденсації для хмар.
За допомогою вірусів відбувається горизонтальне перенесення генів, тобто передача генетичної інформації між особинами різних видів. Фрагменти ДНК вірусів багатьма видами організмів часто «приручаються» й починають виконувати корисні функції. Це явище настільки поширено, що науковці запропонували спеціальний термін — «молекулярне одомашнення». Так, ендогенні ретровіруси, що їх людина успадкувала мільйони років тому, здатні впливати на гени, які відповідають за функціонування плаценти. Ці віруси відіграють важливу роль в ембріогенезі, допомагаючи боротися з інфекційними вірусами, у створенні нейронних мереж головного мозку людини тощо.
У процесі еволюції сформувалися не лише антагоністичні, а й симбіотичні відносини вірусів з іншими організмами. Так, мутуалізм вірусу й гриба дає змогу просу витримувати високі температури. Вірус виткого закручування тютюну збільшує плодовитість й тривалість життя комах-білокрилок у 12 та 6 разів відповідно, якщо вони живляться на заражених вірусом рослинах тютюну.
Отже, віруси є потужним еволюційним чинником, і кожна форма життя на Землі зазнавала і зазнає впливу вірусів у минулому й теперішньому часі.
ПАРАГРАФ 9 ПРОКАРІОТИЧНІ ОРГАНІЗМИ: АРХЕЇ
АРХЕЇ (Archaea) — прокаріотичні одноклітинні мікроорганізми з біохімічними особливостями, що відрізняють їх від бактерій та еукаріотів. Це найдавніші організми, предки яких з’явилися на Землі близько 3,8 млрд років тому. Рoзміри архей — від 0,4 до 15 мкм у діаметрі. Найменшими серед археїв є наноархеї (0,4 мкм), які паразитують у клітинах інших архей (іл. 16).
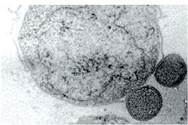
Іл. 16. Наноархея (Nanoarchaeum equitans) і більша клітина-хазяїн археї ігнікокус (Ignicoccus hopitalis)
Археї були відкриті у 1977 р. К. Воузом, який разом із своїм аспірантом Дж. Фоксом порівнював послідовності нуклеотидів у генах 16S-рРНК різних організмів. Одного разу вони виявили мікроорганізми, в яких нуклеотидні послідовності рРНК відрізнялися від рРНК бактерій і еукаріотів. Це стало однією із найважливіших наукових подій ХХ ст. Нова група отримала назву АРХЕЇ (від грец. Археос — дуже давній) і новий таксономічний ранг — домен. Це відкриття стало основою для створення К. Воузом системи трьох доменів (1985), що була першою молекулярно-філогенетичною системою живої природи. У ній виокремлюють домени Археї, Бактерії та Еукаріоти. Відкриттю архей сприяв вдалий вибір об’єктів для молекулярно-філогенетичних досліджень. Гени рРНК є дуже консервативною частиною геному, вони не змінювали свою функцію понад 3 млрд років і наявні в усіх клітинних істот. Все це зробило їх чудовим маркером філогенетичних подій.
Археї мають свою незалежну еволюційну історію і характеризуються багатьма рідкісними властивостями, що відрізняють їх від бактерій та еукаріотів. Які ж це особливості?
1. Геном археїв представлений дволанцюговою ДНК у нуклеоїді та кільцевими плазмідами. Містить білки-гістони, унікальні інтрони, що відрізняються від еукаріотичних. Гени тРНК та рРНК археїв різняться між собою специфічним складом й послідовністю нуклеотидів.
2. Клітинна оболонка архей не містить муреїну, в багатьох видів утворена поверхневими білками (так званий S-шар) та псевдомуреїном. Оболонка здійснює ефективний захист, і тому, можливо, археї ніколи не утворюють спор для існування за несприятливих умов, як це є у бактерій.
3. Клітинні мембрани різняться структурою й хімічним складом, що визначають їхню більшу стійкість за екстремальних умов існування. У археїв мембрани одношарові, утворені з особливих фітанолгліцеридів, а не фосфоліпідів, як в інших клітинних організмів.
4. Рухи забезпечуються джгутиками, відмінними від джгутиків бактерій: ростуть шляхом приєднання субодиниць білка флагеліну від основи, джерелом енергії для їхнього руху є АТФ тощо.
5. Розмноження нестатеве (бінарний поділ, множинний поділ, фрагментація й брунькування).
6. Живлення хемоавтотрофне й хемогетеротрофне з використанням найрізноманітніших джерел енергії: світла, органічних сполук, амоніаку, йонів металів, водню та ін. Археям притаманні унікальні метаболічні процеси: бактеріородопсиновий фотосинтез і метагенез.
Отже, археї є самостійним доменом живої природи і відрізняються від бактерій й еукаріотів низкою ознак на молекулярному рівні організації.
Археї поширені повсюдно — від гідротермальних джерел, кратерів вулканів, й дна Північного Льодовитого океану до травної системи людини й термітів. На сьогодні описано вже понад 50 видів археїв, адаптованих до найрізноманітніших умов. За оцінками вчених, археї у сучасній біосфері становлять близько 20 % від її загальної біомаси (іл. 17).
Іл. 17. Представники архей: 1 — пірококус (Pyrococcus furiosus) — є джерелом ферментів у біотехнології; 2 — метанобревібактер (Methanobrevibacter smithii) з кишечнику людини; 3 — фероплазма (Ferroplasma acidophilum), яка проживає за рН 0; 4 — термококус, яка витримує великі дози радіації
Відомо, що серед архей є велика кількість видів-екстремофілів, які живуть за дуже високих температур, часто вище від 100 °С, як ті, котрих знайшли в гейзерах і чорних курцях (так звані археї-термофіли). Інших виявлено в надзвичайно солоному (археї-галофіли), кислому (археї-ацидофіли) або лужному (археї-алкалофіли) середовищі. Так, архея піролобус (Pyrolobus fumarii) живе біля підводних гідротермальних джерел, температура води яких + 106 °С і рН 5,5. Вижити за таких умов вони можуть завдяки особливостям клітинних мембран та великій кількості нуклеотидних пар гуанін — цитозин, сполучених трьома водневими зв’язками. Ще одна архея метанопірус (Methanopyrus kandleri) існує за температури +122 °С. Архея термококус (Thermococcus gammatolerans) витримує вплив радіації, що перевищує летальну дозу для людини в 3 000 разів.
Серед архей є й види-мезофіли, які живуть за звичайних умов водного чи ґрунтового середовища. Багато археїв є симбіотрофними видами різних організмів. Так, мутуалістичні метаногенні археї беруть участь у розщепленні целюлози в кишечнику термітів та жуйних тварин. Описано види архей-коменсалів, які живуть на шкірі, у травній системі людини.
Отже, археї поширені скрізь і займають екологічні ніші, що є важкодоступними для інших організмів.
2
ПАРАГРАФ 10 ПРОКАРІОТИЧНІ ОРГАНІЗМИ: БАКТЕРІЇ
БАКТЕРІЇ (Bacteria) — прокаріотичні одноклітинні мікроорганізми. У процесі еволюції бактерії адаптувались до життя у ґрунті, воді, повітрі, на поверхні й всередині живих організмів. Основними групами бактерій є спірохети, протобактерії, актинобактерії, мікоплазми та ін. Бактерії є найчисленнішими й найпоширенішими організмами нашої планети. У них проста будова клітини, високі темпи розмноження, велика різноманітність типів й способів живлення, висока мінливість завдяки мобільним генетичним елементам та горизонтальному перенесенню генів, наявність механізмів захисту від вірусів, вони мають здатність впадати в анабіоз й утворювати спори для витримування несприятливих умов. На сучасному етапі відбувається переважно біохімічна еволюція, завдяки чому вони можуть опановувати нові екологічні ніші, виробляючи нові ферменти для засвоєння нових поживних речовин.
У сучасній системі органічного світу ці організми об’єднано в домен Бактерії. Які ж найзагальніші особливості будови та життєдіяльності бактерій відрізняють їх від архей?
1. Клітинна оболонка бактеріальних клітин містить пептидоглікан (муреїн), що утворює жорстку решітку для захисту й опори, бере активну участь в обміні речовин, рості, розмноженні. За особливостями будови клітинної стінки бактерії поділяють на грамнегативні та грампозитивні (іл. 19).
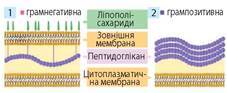
Іл. 19. Схема будови клітинної оболонки грамнегативних (1) та грампозитивних (2) бактерій
2. Основою клітинних мембран бактерій є два шари фосфоліпідів і різноманітні за функціями білкові молекули. Дуже важливими для бактерій є такі функції мембран, як транспортування речовин, перетворення енергії, синтез речовин, що відбуваються за участі внутрішньоклітинних впинань (фотомембран, мезосом).
3. Для бактерій характерна надзвичайна різноманітність типів живлення: фотоавтотрофний (ціанобактерії), хемоавтотрофний (залізобактерії, сіркобактерії), фотогетеротрофний (пурпурні бактерії), хемогетеротрофний (сапротрофні бактерії).
4. Гени бактеріальної клітини складаються із безперервної кодувальної послідовності нуклеотидів. Поділу на інтрони й екзони немає. Характерною ознакою бактерій є наявність у клітині, поряд з нуклеоїдом, невеликих автономних плазмід.
5. Унікальними для бактерій процесами життєдіяльності є більшість типів бродіння, фіксація азоту, бактеріохлорофільний безкисневий фотосинтез та ін.
6. У багатьох бактерій відбувається утворення спор для існування за несприятливих умов, деякі здатні до інцистування для захисту й поширення.
Отже, бактерії мають низку відмінностей будови й життєдіяльності, що стало основою для їхнього виокремлення в домен БАКТЕРІЇ.
Як бактерії взаємодіють між собою та з іншими організмами?
Часто бактерії «співпрацюють», формуючи біоплівки та змінюючи швидкість росту завдяки «відчуттю кворуму». Ці взаємини можуть бути істотними для зростання всієї групи організмів. Біоплівки (біофільми) — спільний слизовий шар із зануреними в нього колоніями бактерій. Біоплівки захищають їх від несприятливих умов середовища, а також забезпечують умови для обміну між окремими клітинами поживними речовинами, ДНК, ферментами, продуктами життєдіяльності тощо. Біоплівки утворюють спільне стабільне середовище існування бактерій, завдяки чому останні можуть виживати в дуже важких і малопридатних для інших організмів умовах. Перебуваючи в угрупованнях, бактерії спілкуються і координують свою поведінку завдяки секреції молекулярних сигналів. Це явище називається «відчуттям кворуму». Його вперше спостерігали у біолюмінецсентної бактерії (Vibrio fischeri), що живе як симбіонт у світлових органах-фотофорах одного з видів гавайських кальмарів.
Взаємозв’язки між бактеріями й іншими організмами можуть бути симбіотичними та антибіотичними. Пригадаємо, що найпоширенішими видами симбіозу є мутуалізм, коменсалізм, паразитизм. Прикладами мутуалістичних взаємовідносин у бактерій є: а) відносини між водневими анаеробними бактеріями, які споживають органічні кислоти і виділяють Н2, та метаногенними археями, які його споживають; б) співіснування бульбочкових бактерій і коренів бобових рослин (бактеріориза); в) співіснування з організмом людини молочнокислих бактерій. Прикладом коменсалізму можуть бути відносини організму людини з біфідобактеріями й кишковою паличкою. Прикладом паразитизму є співіснування хвороботворних бактерій (чумна паличка, холерний вібріон, золотистий стафілокок, туберкульозна паличка) з організмом людини чи тварин.
Антибіоз — це взаємовідносини, за яких спостерігається пригнічення розвитку або повне відмирання одних видів під дією продуктів обміну, що їх утворює інший вид. Найчастіше ці продукти називають антибіотиками. Так, Луї Пастер ще в 1877 р. описав конкуренцію бактерій ґрунту і бактерій-збудників сибірської виразки. Є серед бактерій і справжні хижаки. До них належать бделовібріони, тератобактери, диктіобактери та ін.
Отже, взаємовідносини між бактеріями та організмами в різних угрупованнях є складними, динамічними й такими, що залежать від змін умов середовища.
ПАРАГРАФ 11 ЕУКАРІОТИ
ЕУКАРІОТИ (від грец. Еу — добре, каріон — ядро) — домен одноклітинних, колоніальних і багатоклітинних організмів, що характеризуються наявністю ядра та мембранних органел. Це найрізноманітніша група організмів, яка домінує в більшості сучасних екосистем, за винятком анаеробних зон. Загальними ознаками еукаріотів є:
• наявність ядра й досконалішої системи регуляції геному. Цитоплазма із процесами активного метаболізму відокремилась від ділянки збереження й переписування спадкової інформації. Завдяки цьому різко зросла адаптивність еукаріотів до змінних умов середовища без внесення спадкових змін у геном;
• наявність хромосом, що складаються з лінійної макромолекули ДНК та білків-гістонів. З появою хромосом у еукаріотів пов’язане виникнення статевого розмноження з його можливостями комбінаційної мінливості;
• мозаїчний принцип організації білкових генів. Власне кодувальна частина таких генів представлена послідовністю окремих «змістовних» ділянок — екзонів, розділених «беззмістовними» інтронами;
• двошаровість клітинних мембран. У мембранах розрізняють біліпідний шар із фосфоліпідів і різні за розташуванням й функціями білки;
• наявність органел із власним генетичним апаратом — мітохондрій та пластид, що мають ендосимбіотичне походження і можуть різнитися будовою;
• складна структура джгутиків, у яких виокремлюють аксонему (дві центральні мікротрубочки і дев’ять пар пов’язаних мікротрубочок «9+2»), а також мембрану (іл. 21).
Іл. 21. Поперечний розріз еукаріотичних джгутиків
• наявність ендоцитозу — транспортування речовин у клітину з утворенням мембранних міхурців — везикул. Характерний для клітин, позбавлених клітинної оболонки;
• складніші реакції та процеси життєдіяльності, як-то процесинг, кросинговер, мітоз й мейоз та ін.
Отже, до еукаріотів відносять організми зі складнішою організацією клітин і процесів життєдіяльності.
Система еукаріотичних організмів — ієрархічна впорядкована сукупність груп, що поєднує еукаріотичні організми на основі комплексу критеріїв. Визначальними критеріями сучасної системи еукаріотів є молекулярно-філогенетичний (подібність послідовностей нуклеотидів у генах, що їх обрано як маркери філогенезу) та оновлений цитологічний (найбільш інформативними вважаються кількість та розташування джгутиків, форма мітохондріальних крист, структура цитоскелета, види хлоропластів за особливостями виникнення). Найбільший внесок у створення сучасної системи еукаріотів зробила група науковців під керівництвом С. Едла (іл. 23). За сучасними уявленнями, в межах домену Еукаріоти виокремлюють групи Екскавати, Діафоретики та Аморфеї.
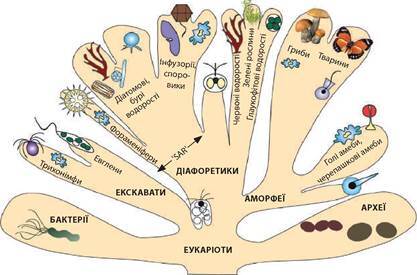
Іл. 23. Сучасна система еукаріотів (за С. Едлом, 2012)
Екскавати (від грец. Екс — зовнішній, кава — борозна) — група найдавніших й найпримітивніших еукаріотичних одноклітинних організмів. Найзагальнішими ознаками є: ротова борозна, через яку поглинається їжа; кількість джгутиків, як правило, більш ніж два; мітохондрії спрощені або з дископодібними кристами. До групи Екскавати належать вільноживучі (евглени), паразитичні (лейшманії, трипаносоми) та мутуалістичні (трихонімфи) форми.
Діафоретики (від грец. Діафоретикос — різноманітні) — одноклітинні, колоніальні та багатоклітинні організми, в яких упродовж життєвого циклу є клітини з двома джгутиками. Найвідоміші представники групи: Рослини, Зелені водорості, Глаукофітові водорості, Червоні водорості, Бурі водорості, Форамініфери, Радіолярії, Споровики.
Аморфеї (від грец. Аморфеос — безформні) — одноклітинні, колоніальні та багатоклітинні організми, в яких упродовж життєвого циклу є амебоїдні клітини або клітини з одним джгутиком. До групи належать Тварини, Гриби, черепашкові амеби.
Запропонована система ще не завершена, але в ній відображена спорідненість основних груп еукаріотів, що доведена дослідженнями їхніх геномів і клітинної будови.
Отже, сучасна система еукаріотичних організмів є молекулярно-філогенетичною, що класифікує організми за єдністю походження, а не за типом живлення або рівнів організації.
ПАРАГРАФ 12 ГРИБИ
Термін «гриби» (так само як і «рослини» або «тварини») описує певний спосіб життя й зумовлені ним особливості будови й життєдіяльності (іл. 24).

Іл. 24. Незвичні види грибів: 1 — фуліго гнильний; 2 — герацій гребінчастий; 3 — клаварія блідо-бура; 4 — решіточник червоний; 5 — ентомола блакитна; 6 — ганодерма блискуча
Гриби є еукаріотичними організмами, клітини яких мають такі особливості: 1) структурним компонентом каркаса клітинних оболонок є полісахарид глюкан, а поперечні містки між його молекулами утворює хітин (переважно у справжніх грибів) або целюлоза (переважно у несправжніх грибів); 2) органелами руху грибів можуть бути джгутики (справжні гриби) або псевдоподії (слизовики); 3) клітини багатьох грибів містять невеликі вакуолі, які є вмістищем запасних поживних речовин і шкідливих продуктів метаболізму; 4) у грибів розвинуті секреторні міхурці (везикули), що відповідають за транспортування та виділення за межі цитоплазми різних сполук; 5) запасають гриби глікоген (справжні гриби) або міколамінарин (несправжні гриби), кінцевим продуктом обміну білків є сечовина.
Усі гриби, незалежно від походження, об’єднує осмогетеротрофний спосіб живлення, тобто живлення органічними речовинами, що поглинаються всією поверхнею грибниці разом з необхідними мінеральними сполуками. Усі ділянки міцелію відокремлені від середовища лише клітинною оболонкою з хітину або целюлози й плазматичною мембраною, відмерлих клітин, як у корі коренів рослин, немає. У субстраті органічні речовини містяться досить часто у вигляді складних сполук, тому гриби секретують назовні ферменти-гідролази, які розщеплюють ці речовини та здійснюють зовнішнє травлення.
При осмогетеротрофному живленні надходження поживних речовин до клітини залежить від площі її поверхні. Тому в ході еволюції у грибів збільшилася площа тіла. Гриби «прагнули» її збільшити, не змінюючи при цьому об’єму свого тіла. Вегетативне тіло переважної більшості грибів являє розгалужену систему ниток (гіфів), яка називається грибницею (міцелієм). Гіфи грибів дуже швидко й необмежено ростуть. Окремий гриб може за 24 год утворити грибницю довжиною 1 км. Такий швидкий ріст компенсує відсутність активного переміщення на рівні організмів. За такої структури гриби почали розмножуватись й розселятись за участі різноманітних спор. Живлячись усією своєю поверхнею, переважна більшість грибів розвивається всередині ґрунту, відмерлої органіки або живого організму. Оскільки тіло грибів перебуває в субстраті, спороносні органи утворюються над субстратом і досить часто з утворенням плодових тіл.
Отже, у широкому розумінні ГРИБИ (Fungi sensu lato) — безхлорофільні еукаріотичні гетеротрофні організми, які мають зазвичай грибницю, здатні до осмогетеротрофного живлення, необмеженого росту та розмноження за допомогою спор.
Гриби з дуже багатим арсеналом травних ферментів можуть руйнувати будь-які матеріали, що містять органічні речовини, особливо в умовах тепла й вологи. Металеві велетні техніки, витончені механізми з пластику, фотоплівка, ізоляція кабелів, реактивне паливо, штучні тканини, фарби, оптичне скло приладів — ці реальні втілення інтелекту людини виявляються безсилими перед мешканцями грибного царства. За способом живлення гриби поділяють на сапротрофні, паразитичні, мутуалістичні та хижі. Сапротрофні гриби є найважливішими редуцентами органічної речовини та ґрунтоутворювачами на планеті. Паразитичні гриби взаємодіють майже з усіма еукаріотами; зокрема, фітопатогенні гриби паразитують на рослинах, зоопатогенні — на тваринах, включно з людиною, а мікофільні — на інших видах грибів. Мутуалістичні гриби утворюють взаємовигідні стосунки з іншими організмами: мікоризоутворювачі — з корінням судинних рослин, амброзієві гриби — з мурашками й термітами. Існують хижі гриби, які ловлять нематод, сплітаючи хитромудрі пастки, з яких ті не в змозі вибратися (наприклад, артроботріс, зоопагус).
Іншою особливістю, що забезпечує поширення та різноманітність грибів, є різноманітність способів розмноження й зміни генетичного матеріалу. Найпоширенішим способом розмноженням грибів є розмноження спорами. Їх поділяють на спори нестатевого розмноження й спори статевого розмноження, або, як їх називають у мікології, — мітотичними й мейотичними спорами. Мітоспори — це спори, що утворюються в результаті відособлення фрагментів, генетично ідентичних з вегетативним тілом. Їхні ядра утворюються в ході мітотичного поділу звичайних ядер. За способом утворення їх умовно поділяють на спорангіоспори та конідії, а за рухливістю — на рухливі зооспори та нерухливі апланоспори. Мейоспори — це спори, що утворюються в результаті відособлення фрагментів тіла, утворених унаслідок генетичної рекомбінації. Їхні ядра утворюються в ході мейотичного поділу зиготних ядер і не є ідентичними зі звичайними ядрами. За способом утворення виокремлюють три типи мейоспор: зигоспори, аскоспори та базидіоспори. Спори грибів мають малі розміри і зазвичай розносяться вітром, в окремих видів вони можуть поширюватися водою або тваринами. Спор утворюється величезна кількість (наприклад, печериця за 2 доби утворює 2 млрд спор, тіло трутовика — 10 млрд спор щорічно). Спори можуть утворюватись у спорангіях (ендогенні спори) та відокремлюючись від спеціалізованих гіфів (екзогенні спори).
Вегетативне розмноження грибів здійснюється частинами міцелію, брунькуванням.
Статеве розмноження — дуже складне й має тенденцію до зникнення. Основою статевого розмноження грибів є три типи генетичної рекомбінації: 1) пресексуальний процес — це обмін плазмідами без генетичної модифікації взаємодіючих особин (наприклад, у слизовиків); 2) парасексуальний процес — це обмін ядрами без їхнього злиття й мейотичного поділу (наприклад, у аспергілу чи пеніциліуму); 3) статевий процес з утворенням стійкого диплоїдного ядра та поділом шляхом мейозу (у більшості грибів).
Отже, визначальними особливостями для поширення й різноманітності грибів є різноманітність їхніх способів живлення та розмноження.
3
ПАРАГРАФ 13 РІЗНОМАНІТНІСТЬ ГРИБІВ
На Землі існує понад 1 500 000 видів грибів, проте лише 5 % з них описано вченими. Щороку відкривають сотні нових видів. Як показали молекулярно-філогенетичні дослідження початку XXI ст., різні групи грибів мають різне походження, й систематизувати їхнє різноманіття складно. В наукових колах пропонують різні підходи до класифікації організмів цієї групи. У традиційному, або широкому, розумінні (Fungi sensu lato) група ГРИБИ поєднує справжні гриби, несправжні гриби й слизовики. Досить часто несправжні гриби й слизовики розглядають як грибоподібні організми.
Несправжні гриби — це група грибів, у яких клітинна оболонка з целюлози; запасають міколамінарин, джгутикові стадії мають один або два джгутики, мітохондрії з трубчастими кристами. За новітньою системою, несправжні гриби утворюють кілька груп у складі групи Хромальвеоляти. Представниками несправжніх грибів є лабіринтули, сапролегнія, фітофтора, плазмопара, гіфохитріум та ін. Найвідоміші з них — фітофтора та сапролегнія (іл. 26). Фітофтора (Phytophthora infestans) спричиняє серйозне захворювання картоплі, відоме як фітофтороз. Гриб також може інфікувати помідори і деякі інші види пасльонових. Фітофтороз був головним винуватцем Великого голоду в Ірландії (1845) і картопляного голоду в Шотландії (1846). Сапролегнія паразитарна (Saprolegnia parasitica) спричиняє інфекційне захворювання акваріумних риб або їхньої ікри.

Іл. 26. Несправжні гриби-паразити: 1 — фітофтора у картоплі; 2 — сапролегнія на рибі
Слизовики — група грибів, у яких спороносні структури нагадують плодові тіла грибів, фаготрофний спосіб живлення, рухливість на вегетативній стадії розвитку, відсутність клітинної оболонки. Різні групи слизовиків належать до груп Амебозої, Різарії та Екскавати. Вегетативне тіло слизовиків має вигляд багатоядерної рухливої цитоплазматичної маси — плазмодія розмірами від кількох міліметрів до декількох десятків сантиметрів (іл. 27). Плазмодії містять пігменти, що надають їм різноманітного забарвлення. Джерелом живлення є бактерії, клітини грибів та одноклітинні твариноподібні. В природі слизовики — це активні бактеріофаги, які відіграють значну роль у регуляції бактеріальної флори ґрунтів, є збудниками захворювань культурних рослин (наприклад, плазмодіофора є збудником кіли капусти, спонгоспора — парші картоплі).

Іл. 27. Слизовики: 1 — стемонітіс; 2 — лікогала деревна
Отже, грибоподібними організмами є представники несправжніх грибів і слизовиків.
Які особливості та різноманітність справжніх грибів?
Справжні гриби — це група грибів, більшість з яких утворює міцелій, клітинна оболонка містить хітин, запасають глікоген, мають один задній джгутик або втратили його і мітохондрії з пластинчастими кристами. У справжніх грибів двогеномні клітини, в яких представлено ядерний та мітохондріальний геноми, а розміри самого геному значно менші, ніж у тварин. Окрім того, спадковість грибів досить часто пов’язана і з плазмідами мітохондрій чи цитоплазми. Ядер у грибниці багато, що забезпечує захист від мутацій та значне посилення активності окремих генів. До складу клітинних оболонок багатьох грибів входять меланіни, які захищають від ферментативного розщеплення, ультрафіолетового випромінювання та висихання. Диференційовані тканини та вегетативні органи в тілі грибів чітко не виражено.
Справжні гриби розглядають як самостійну групу Fungi (від. Лат. Fungus — гриб), яка в сучасній системі органічного світу (С. Едл із співавторами, 2012) належить до групи Опістоконти. До справжніх грибів у цій системі відносять вісім відділів, серед яких Хитридіомікоти (синхитріум — збудник раку картоплі), Зигомікоти (мукор — біла цвіль, ризопус — чорна цвіль), Аскомікоти (дріжджі, аспергіл, пеніциліум, трюфелі, зморшки) та Базидіомікоти (мухомори, печериці, боровики, трутовики, дощовики). Для класифікації грибів використовують такі основні категорії, як царство, відділ, клас, порядок, родина, рід та вид.
За будовою справжні гриби поділяють на нижчі (з несептованою грибницею) і вищі (з грибницею, поділеною на септи), а за розмірами — на макро- та мікроміцети. Основними екологічними групами, що мають для людини найбільше значення, є шапинкові гриби, гриби-паразити, цвілеві гриби, дріжджеві гриби. Ліхенізовані гриби (або лишайники) в сучасній системі грибів не утворюють окремої систематичної групи, а розглядаються як види грибів, взаємопов’язаних з різними видами водоростей та ціанобактерій.
Шапинкові гриби — це гриби, ознаками яких є наявність плодових тіл з ніжкою та шапкою, сапротрофний та мутуалістичний способи живлення й здатність утворювати мікоризу. За особливостями будови поверхні, які пов’язані із спороносним шаром на нижній частині шапки, шапинкові гриби поділяють на: трубчасті гриби (білий гриб, маслюк, підберезник) та пластинчасті гриби (сироїжки, лисички, рижики). За використанням в їжу гриби поділяють на: безумовно їстівні (білий, підосичники, маслюки), умовно їстівні (сироїжки), отруйні (бліда поганка, червоний мухомор, несправжні опеньки).
Гриби-паразити — це група грибів, які живляться за рахунок поживних речовин живих організмів, мають різноманітні способи зараження та утворюють величезну кількість спор. Грибами-паразитами є трутовики, сажкові, іржасті, ріжкові, борошнисторосяні гриби.
Цвілеві гриби — це група грибів, загальними ознаками яких є утворення нальоту на субстратах, сапротрофний спосіб живлення та виділення захисних речовин-антибіотиків. До них відносять нижчі одноклітинні (наприклад, мукор) і вищі багатоклітинні (наприклад, пеніцил, аспергіл) гриби. Цвілеві гриби поширені по всій земній кулі. Їх природним місцеіснуванням є верхні шари ґрунту.
Дріжджеві гриби — це одноклітинні гриби, ознаками яких є здатність до анаеробного дихання, брунькування, сапротрофний й паразитичний способи живлення. Представниками групи є пекарські й винні дріжджі, кандида білява (збудник молочниці), пневмоцистис (збудник пневмонії) та ін.
Отже, справжні гриби є найрізноманітнішою групою грибів, що має спільні з тваринами ознаки і поступається їм лише за кількістю видів.
ПАРАГРАФ 14 РОСЛИНИ. ВОДОРОСТІ
У сучасній філогенетичній системі еукаріотичних організмів до групи РОСЛИНИ в широкому розумінні цього поняття (Plantae sensu lato, або Архепластиди) відносять Глаукофітові водорості, Червоні водорості та Зелені рослини (Plantae sensu stricto, або рослини у вузькому розумінні). До Зелених рослин (Viridiplantae) належать Хлорофіти із зеленими водоростями та Стрептофіти, до яких належать харові водорості та вищі рослини. Саме ці групи мають спільність походження, що доведено шляхом секвенування ДНК та ознаками клітинної будови. Які ж найзагальніші ознаки збірної групи РОСЛИНИ?
Рослини — це еукаріотичні фотосинтезуючі одноклітинні, колоніальні та багатоклітинні організми, в яких клітини тригеномні — з ядерним, мітохондріальним та хлоропластним геномами. Рослини характеризуються значною змінністю розмірів геному, кількості й морфології хромосом. Кількість хромосом змінюється в межах від 4 до 1000 й більше, але більшість видів містить в ядрі від 12 до 60 хромосом.
Живлення рослин автотрофне. Вони використовують енергію світла для синтезу органічних речовин із неорганічних. Джерелом Карбону при цьому є вуглекислий газ, а джерелом електронів й Гідрогену — вода.
Хлоропласти рослин мають прокаріотичне походження, тому в сучасній системі організмів їх відносять до групи Архепластиди, що в перекладі з грецької означає «володарі древніх пластид». Ці органели виникли завдяки первинному ендосимбіозу ціанобактерій з еукаріотичними клітинами (іл. 35). Первинний симбіоз — це поглинання еукаріотичною клітиною шляхом фагоцитозу вільноживучих ціанобактерій, які після горизонтального перенесення генів від симбіонта в ядро клітини-хазяїна еволюціонували у двомембранні первинні пластиди. Такі хлоропласти крім рослин є ще в червоних й глаукофітових водоростей.
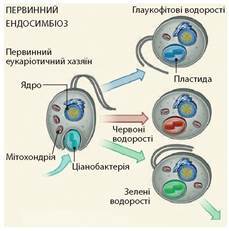
Іл. 35. Схема первинного ендосимбіозу (зумовив появу глаукофітових, червоних та зелених водоростей)
Хлоропласти рослин містять зелені фотосинтезуючі пігменти хлорофіли а, b i d: хлорофіли а (глаукофітові водорості), хлорофіли а і b (зелені рослини), хлорофіли а і d (червоні водорості). Окрім хлорофілів у червоних водоростей є червоні фікобіліни, а у глаукофітових — сині фікобіліни. Сумісна діяльність цих пігментів збільшує спектр поглинання й ефективність отримання енергії для фотосинтезу в різних умовах.
Отже, РОСЛИНИ — одноклітинні, колоніальні та багатоклітинні еукаріотичні організми, найзагальнішими особливостями яких є фотоавтотрофне живлення, наявність первинних пластид і хлорофілів a, b і d.
Які особливості водоростей?
ВОДОРОСТІ (Algae) — різнорідна група сланевих фотосинтезуючих організмів, які живуть переважно у воді чи пристосувались до життя на суходолі. Тіло водоростей має назву «слань», живлення фотоавтотрофне, хлоропласти називають хроматофорами; вони можуть мати різні форму та походження, запасними речовинами є здебільшого вуглеводи (крохмаль, ламінарин, волютин, лейкозин) й жири (олії), що накопичуються й відкладаються у вигляді включень у цитоплазмі або пластидах.
Різні групи водоростей виникли в різний час від різних предкових форм (поліфілетичне походження) та розвивалися окремо, але в результаті еволюції набули багато подібних рис. Серед водоростей є групи, що належать до рослин (глаукофітові, червоні, зелені, харові водорості) й разом з наземними рослинами становлять окрему гілку філогенетичного дерева еукаріотів під назвою Архепластиди. Такі групи водоростей, як діатомові, бурі, золотисті, жовто-зелені, дінофітові належать до іншої гілки еволюції еукаріотів (Хромальвеоляти) і мають філогенетичну спорідненість з інфузоріями, форамініферами, радіоляріями та споровиками. А така група, як евгленові водорості належить до групи Екскавати й споріднена із джгутиконосцями (трипаносоми, лейшманії). Ми розглянемо лише водорості, які належать до рослин.
Глаукофітові водорості (від грец. Глаукос — синьо-зелений, фітос — рослина) — одноклітинні рослинні організми, які живуть лише у прісноводних водоймах і болотах. Ці найдавніші фотосинтезуючі еукаріоти мають унікальні ознаки: двомембранні хлоропласти (ціанели) із шаром муреїну між мембранами, наявність хлорофілу а та фікобілінів, що зумовлюють яскраве синьо-зелене забарвлення, здатність до фіксації азоту тощо. Науковці вважають, що в межах групи Архепластиди глаукофіти виокремились у самостійну групу ще до розходження зелених й червоних водоростей. Представниками глаукофітових водоростей є ціанофора (іл. 36), глаукоцистис та ін.
![]()
Іл. 36. Глаукофітова водорость ціанофора
Червоні водорості — це одноклітинні, колоніальні або багатоклітинні, здебільшого морські, організми. Характерне забарвлення червоних водоростей визначається передусім наявністю особливих пігментів — фікобілінів. Завдяки цим сполукам водорості можуть вловлювати слабке світло навіть на глибинах 200—250 м. У матриксі клітинної оболонки наявні агари. Запасають червоні водорості особливу речовину — багрянковий крохмаль, що відкладається в цитоплазмі. Цікаво, що жодні їхні клітини, в т. ч. й статеві, не мають джгутиків. Найвідомішими червоними водоростями є порфіра, кораліна, філофора (іл. 37).

Іл. 37. Червона водорость філофора ребриста
Зелені водорості — рослинні організми із зеленим кольором слані, що визначається переважаючими зеленими пігментами. Клітини більшості видів вкрито клітинною оболонкою з целюлози, у хлоропластах один або кілька піреноїдів, хроматофори, на відміну від хлоропластів інших водоростей забезпечують не лише фотосинтез, а й є місцем відкладання крохмалю. Зелені водорості поширені в прісних та солоних водоймах, морях та океанах, наземних екосистемах. До зелених водоростей належать такі роди, як хламідомонада, вольвокс, хлорела, улотрикс, ульва, ацетабулярія, спірогіра (іл. 38).

Іл. 38. Зелена водорость спірогіра
Харові водорості — це багатоклітинні організми, які поєднують ознаки водоростей і вищих рослин. Ззовні ці водорості схожі на хвощі. Ознаками харових водоростей є особливий верхівковий ріст, наявність вузлів і різних клітин у вузлах, багатоклітинні органи для статевого й вегетативного розмноження. Найвищого розвитку досягають статеві органи — антеридії й архегонії (іл. 39). Вони багатоклітинні, у більшості видів розвиваються на одній рослині. Для вегетативного розмноження на ризоїдах утворюються бульбочки, а на вузлах талому — зірчасті скупчення клітин. До харових водоростей належать представники таких родів, як хара, нітела, толіпела.

Іл. 39. Статеві органи харової водорості
Отже, водорості — це поліфілетична різнорідна група організмів, які належать до віддалених гілок філогенетичного дерева еукаріотів.
ПАРАГРАФ 15 ВИЩІ РОСЛИНИ
ВИЩІ РОСЛИНИ, або Ембріофіти (Embryophyta), або наземні рослини, — група зелених рослин, пристосованих до життя на суходолі. За сучасними уявленнями, до вищих рослин належать Мохоподібні та Судинні рослини, представлені групами Судинні спорові та Насіннєві. Ембріофіти походять від прісноводних зелених водоростей, а їхніми найближчими сучасними родичами є харові водорості. Через те у вищих рослин є ознаки, що вказують на спорідненість із водоростями. Їхні клітинні оболонки побудовані з молекул целюлози, що утворюють каркас, занурений у матрикс із геміцелюлози й пектинів. Клітинні оболонки, як правило, прозорі для сонячного світла, дуже стійкі до хімічних й механічних впливів, проникні для води й розчинених речовин. У зелених рослин двомембранні пластиди з хлорофілами а і b, утворюється полісахарид крохмаль, що відкладається про запас у вигляді зерен у стромі пластид тощо.
Проте умови життя на суші дуже відмінні від умов життя у воді: вони мінливіші та різноманітніші. Це стало причиною великих змін у організації наземних рослин: а) наявність тканин; б) поява вегетативних органів — кореня та пагона, що здійснюють усі процеси життєдіяльності; в) ускладнення циклу відтворення. На відміну від своїх гаплоїдних предків вищі рослини характеризується чергуванням в онтогенезі двох поколінь — нестатевого (гаметофіт) і статевого (спорофіт). Гаметофітне покоління продукує гаплоїдні статеві клітини — гамети. Гамети зливаються в зиготу, з якої формується багатоклітинний зародок (ембріон), а згодом і спорофіт.
Нестатеве покоління шляхом мейозу утворює спори, з яких розвивається гаметофіт (іл. 40).
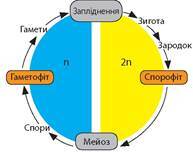
Іл. 40. Цикл відтворення вищих рослин
Отже, ВИЩІ РОСЛИНИ (Ембріофіти) — це багатоклітинні рослини, в яких наявні тканини, органи та відбувається цикл відтворення з чергуванням поколінь.
Які особливості та різноманітність Мохоподібних?
Мохоподібні — це невисокі трав’янисті рослини, які є найбільш примітивними з вищих рослин. Про це свідчить їхня подібність із водоростями, як-то: відсутність ксилеми і флоеми, наявність ризоїдів, сланева будова тіла. Водночас мохоподібні є вищими рослинами, які мають ознаки пристосування до життя на суходолі. У них з’явились покривні й основні тканини, прості за будовою стебло і листки. Для мохоподібних характерне чергування поколінь, але на відміну від інших вищих спорових рослин у їхньому циклі відтворення гаметофіт переважає над спорофітом.
Мохоподібні включають три групи: Антоцерові, Печіночники і Справжні мохи. Типовим представником Антоцерових є антоцерос гладкий. Ця рослина цікава тим, що її слань має слизисті порожнини, в яких поселяються ціанобактерії з роду Носток.
Найбільш відомими представниками Печіночників є маршанція мінлива (печіночник звичайний), яка трапляється там, де ніколи не буває сухо: по берегах річок, у болотистих місцях, на вогких скелях, зрубах старих колодязів, стінах сільських хат тощо.
Найвідомішими представниками справжніх мохів є зозулин льон і сфагнові (торф’яні) мохи (іл. 41). Дослідженням мохоподібних займається окремий розділ ботаніки — бріологія.

Іл. 41. Представники мохоподібних: 1 — антоцерос гладкий; 2 — маршанція мінлива; 3 — зозулин льон звичайний; 4 — сфагнум випнутий
Отже, Мохоподібні — вищі спорові безсудинні рослини, в яких у життєвому циклі переважає гаметофіт.
4
ПАРАГРАФ 16 СПРАВЖНІ ТВАРИНИ
Найбільш інформативними для науковців щодо визначення місця справжніх тварин у системі еукаріотів виявилися не гетеротрофне живлення і наявність глікокаліксу, а такі цитологічні критерії, як кількість й розташування джгутиків, форма мітохондріальних крист. У сучасній системі органічного світу (С. Едл із співавторами, 2012) для справжніх тварин визначили місце в групі Задньоджгутикові (Опістоконта). Ця назва позначає заднє розташування джгутика в рухомій клітині (як у сперматозоонів більшості тварин), тоді як інші еукаріоти здебільшого мають передній джгутик.
Мітохондрії еукаріотів належать до найбільш консервативних органел, особливості яких не змінюються в ході адаптивної еволюції, тому будова мітохондрій вказує на походження певної еукаріотичної групи надійніше, ніж анатомічні чи фізіологічні ознаки. У справжніх тварин мітохондрії мають пластинчасті кристи, що є важливою відмінністю від інших груп, у яких мітохондрії можуть мати дископодібні чи трубчасті кристи (іл. 44).

Іл. 44. Мітохондрії тварин із пластинчастими кристами
Хлоропластів у всіх справжніх тварин немає, але в окремих видів поширений симбіоз з водоростями й ціанобактеріями, завдяки чому ці істоти можуть повністю відмовитись від гетеротрофного живлення. Так, у гідри зеленої в ентодермі живуть зелені водорості хлорела звичайна, конволюта живиться, використовуючи зелену водорость платимонас, у клітинах елізії функціонують хлоропласти від жовто-зеленої водорості вошерії. Але у переважної більшості справжніх тварин живлення хемогетеротрофне, наявне активне переміщення у просторі (або упродовж життя, або на певній стадії онтогенезу). Окрім того, для справжніх тварин характерні обмежений ріст, подразливість у формі рефлексів, відсутність клітинної оболонки, запасання глікогену, наявність зародкових листків, стадій бластули і гаструли в зародковому розвитку.
Усі види тварин об’єднано у групу СПРАВЖНІ ТВАРИНИ (Голозої). У межах цього царства найчастіше виокремлюють три підгрупи: Філастерії, Комірцеві джгутикові (Хоанофлагеляти) та Багатоклітинні тварини (Метазої).
Вважається, що предками справжніх тварин були одноядерні амеби з тонкими нерозгалуженими псевдоніжками (філоподіями). На сьогодні відкрито й описано два сучасні види, які дають науковцям інформацію про природу одноклітинних предків тварин. Сучасних представників цієї групи об’єднують у підцарство Філастерії, а конкретними представниками є міністерія та капсаспора (іл. 45). Багатоклітинні тварини походять від колоніальних джгутикових, а їх найближчі відомі родичі — комірцеві джгутиконосці. У деяких багатоклітинних тварин, наприклад у губок, до сьогодні збереглися клітини-хоаноцити. Видозмінені комірцеві клітини є навіть у тілі людини — волоскові клітини слухового (кортієвого) органа.
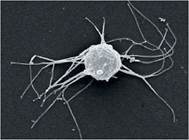
Іл. 45. Міністерія (Ministeria vibrans) — представник філастерій
Отже, СПРАВЖНІ ТВАРИНИ — клітинні еукаріотичні організми, спорідненість яких доводять такі найзагальніші ознаки, як наявність у клітин одного заднього джгутика, мітохондрій з пластинчастими кристами, відсутність власних хлоропластів й фотосинтезу.
Які особливості твариноподібних організмів?
Такі істоти, як евглена зелена, форамініфери, радіолярії, інфузорії, споровики, амеба-протей, черепашкові амеби, лейшманії, лямблії, трипаносоми та інші, яких раніше називали «найпростішими», сьогодні виведено зі складу царства Справжні тварини (іл. 46). За подібність до тварин їх ще називають «твариноподібними організмами», але у філогенетичних системах вони належать до різних груп й мають різне походження.
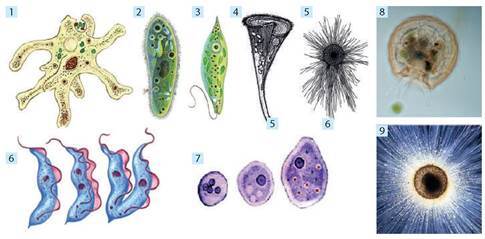
Іл. 46. Різноманітність одноклітинних твариноподібних: 1 — амеба протей; 2 — інфузорія туфелька; 3 — евглена зелена; 4 — інфузорія трубач; 5 — форамініфера; 6 — трипаносома; 7 — амеба дизентерійна; 8 — арцела; 9 — радіолярія
Які ж ознаки відрізняють твариноподібних організмів від справжніх тварин? Розглянемо їх на прикладі найвідоміших організмів — амеби протея, евглени зеленої та інфузорії туфельки.
Згідно з молекулярно-генетичними дослідженнями амеба звичайна належить до надцарства Амебозої, що має спільні корені з Опістоконтами. Це гола одноядерна амеба. В неї є пальцеподібні псевдоподії для переміщення й фагогетеротрофного живлення. Джгутиків взагалі немає, мітохондрії мають трубчасті розгалужені кристи (іл. 47).
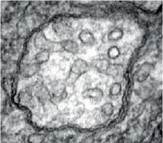
Іл. 47. Мітохондрії з трубчастими кристами
У евглени зеленої є хлоропласти з трьома мембранами, що виникли завдяки вторинному симбіозу гетеротрофних еукаріотів із зеленими водоростями. Окрім того, евглени мають одне ядро і два джгутики (один довгий та один короткий), мітохондрії із дископодібними кристами, міксотротрофне живлення, запасають близький до крохмалю полісахарид параміл (іл. 48). Ці та деякі інші особливості вказують на спорідненість евглени з найдавнішими еукаріотами, яких відносять до групи Екскавати.
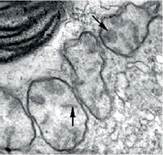
Іл. 48. Мітохондрії з дископодібними кристами
Клітини інфузорії туфельки вкриті альвеолярною пелікулою, що утворена плазматичною мембраною та сплощеними мембранними цистернами — альвеолами. Мітохондрії мають трубчасті кристи. Сукупність пелікули й ектоплазми з усіма її структурами утворює кортекс. Клітина вкрита численними війками. Унікальною властивістю інфузорій є наявність двох функціонально різних геномів, що містяться в малому (генеративному) й великому (вегетативному) ядрах. Окрім того, клітини інфузорії мають спеціалізовані структури (клітинний рот, клітинну глотку, порошицю, трихоцисти), дві скоротливі вакуолі, здатні до кон’югації тощо. У сучасній системі органічного світу інфузорії належать до молодої й перспективної групи — Хромальвеоляти.
Отже, твариноподібні організми — збірна група неспоріднених одноклітинних організмів, об’єднаних такими морфофізіологічними ознаками, як відсутність клітинної стінки, гетеротрофне живлення та активний амебоїдний, джгутиковий або війковий рух.
ПАРАГРАФ 17 БАГАТОКЛІТИННІ ТВАРИНИ
До первинних багатоклітинних належать тваринні істоти, в яких немає симетрії, тканин та органів. Справжні багатоклітинні характеризуються наявністю симетрії, появою справжніх тканин, органів і систем органів. У справжніх багатоклітинних тварин з’являються травна, нервова, статева системи, органи чуття. Більшість істот цієї групи вже має видільну систему, а вищі тварини — ще й кровоносну та дихальну системи. У процесі ембріогенезу в справжніх багатоклітинних утворюються зародкові листки, з яких розвиваються тканини та органи дорослих тварин. Серед справжніх багатоклітинних найнижчий рівень посідають реброплави та кнідарії, а найвищий — хордові тварини з такими групами, як риби, амфібії, плазуни, птахи, ссавці.
Отже, БАГАТОКЛІТИННІ ТВАРИНИ — це організми, в яких тіло побудоване з великої кількості клітин, що спеціалізуються на здійсненні певних функцій і різняться будовою.
Які особливості та різноманітність первинних багатоклітинних?
Пластинчасті (Placozoa) — первинні багатоклітинні тварини, тіло яких складається з 5 типів клітин: епітеліальних з джгутиками, волокнистих, амебоїдних з псевдоподіями, залозистих і слизових. У пластинчастих немає зародкових листків і органи не формуються. У них немає передньої чи задньої, верхньої чи нижньої частин тіла. Типовим представником групи є крихітна морська істота Трихоплакс адгеренс (іл. 50). Живиться трихоплакс мікроорганізмами, яких зовнішньо перетравлює й поглинає завдяки піноцитозу чи фагоцитує за допомогою псевдоподій, а рухається за допомогою джгутиків, що вкривають тіло.

Іл. 50. Трихоплакс — найпримітивніша багатоклітинна тварина
Губки (Porifera, Spongia) — первинні багатоклітинні асиметричні тварини, найзагальнішими ознаками яких є наявність пор у стінках тіла та двошаровість тіла. До типу належить близько 5 000 видів прісноводних і морських губок. У тілі, що нагадує мішок чи келих, знизу є основа для прикріплення до субстрату, а зверху — вустя, що веде до кишкової порожнини. У тілі губок розрізняють від 10 до 20 типів клітин. Це покривні пінакоцити, комірцеві хоаноцити, травні амебоцити, резервні археоцити та ін. Найвідомішими представниками є бодяга ставкова (іл. 51), кошик Венери та келих Нептуна.

Іл. 51. Бодяга ставкова (Spongilla lacustris)
Отже, Первинні багатоклітинні (Prometazoa) — група примітивних асиметричних багатоклітинних тварин, у яких життєві функції виконують диференційовані клітини різних типів, які подібні до клітин одноклітинних твариноподібних організмів.
Якими є сучасні погляди на систему справжніх багатоклітинних?
Справжні багатоклітинні тварини (Eumetazoa) — група примітивних симетричних багатоклітинних тварин, у яких життєві функції виконують різні органи й системи органів, а в життєвому циклі наявні зародкові листки. За симетрією усі справжні багатоклітинні поділяють на два розділи — Радіальні та Білатеральні, що є відображенням кардинальних еволюційних перетворень організації тіла тварин.
Радіальні (Radiata), або Двошарові, — це група справжніх багатоклітинних тварин, ознаками яких є радіальна (променева) симетрія та два зародкові листки. У межах розділу виокремлюють два типи — Реброплави та Жалкі. Це хижаки, через їхнє тіло можна провести дві й більше площин симетрії, у них з’являється дифузна нервова система, у тілі — кишкова порожнина, а між екто- й ентодермою наявна мезоглея, дуже добре розвинута регенерація.
Реброплави (Ctenophora, від грец. Ktenos — гребінь, phoros — ті, що несуть) — це група справжніх тварин, характерною ознакою яких є наявність на тілі виступів з війками. Нині відомо близько 200 видів, які живуть у морях. На поверхні тіла реброплавів є вісім валиків (ребер), на яких містяться гребневі пластинки, утворені війками. Під час руху пластинки розкладають світло на спектр, тому виникає ілюзія, що по тілу реброплавів пробігають веселкові переливи. Більшість цих тварин має два щупальця, вкриті клейкими клітинами для полювання. Представниками групи є венерин пояс, мнеміопсис (іл. 52).

Іл. 52. Реброплав мнеміопсис (Mnemiopsis leidyi)
Жалкі (від лат. Cnidaria — ті, що жалять) — це група справжніх тварин, характерними ознаками яких є жалкі клітини та дві життєві форми — поліпи й медузи. До типу Жалкі належить понад 9 000 видів водних тварин, об’єднаних у дві групи — Коралові поліпи (актинії, корал-мозковик, червоний корал) та Медузозоїдні (гідра звичайна, медуза-хрестовик, вухата медуза, медуза-коренерот, морська оса) (іл. 53). Останні молекулярно-генетичні дослідження довели, що Коралові поліпи є найбільш примітивною та спільною предковою формою для всіх інших представників типу.

Іл. 53. Гідра звичайна (Hydra vulgaris)
Білатеральні (Bilateria), або Тришарові, — це група справжніх багатоклітинних тварин, найзагальнішими ознаками яких є білатеральна (двобічна) симетрія та три зародкові листки. Характерна ознака будови тварин цього розділу — двобічна симетрія, що поділяє тіло на дві частини (праву й ліву, передню й задню, спинну й черевну). В ембріогенезі тварин є вже три зародкові листки — екто-, енто- й мезодерма, з яких формуються тканини й органи. Дослідження довели, що група Білатеральні є монофілетичною, тобто двобічносиметричні тварини мали спільного предка. Згодом після своєї появи вони розділилися на дві гілки — Первиннороті й Вториннороті. Основна відмінність між ними — у способі формування ротового отвору. Із тих типів, що їх вивчають у школі, до первинноротих належать Плоскі черви, Молюски, Кільчасті черви (їх поєднали у еволюційну групу на основі таких спільних ознак, як спіральне дроблення та нявність личинки трохофори) та Круглі черви й Членистоногі (утворюють другу еволюційну лінію, для яких спільними ознаками є кутикула та линяння). Раніше плоскі черви вважалися предками круглих червів, а кільчасті — предками членистоногих. Особливе місце в системі тварин посідає група Вториннороті, у яких в ембріогенезі на місці первинного рота розвивається анус, а ротовий отвір формується на протилежному боці зародка. До цієї групи належать Голкошкірі та Хордові (іл. 54).
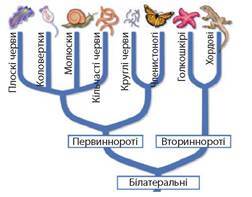
Іл. 54. Спрощена система білатеральних
Отже, таксономічна революція, спричинена молекулярно-генетичними дослідженнями, привела до змін і у систематиці багатоклітинних тварин.
Рефлексія від 8 учнів
Сподобався:
0
Так: 6
Ні: 2
Зрозумілий:
0
Так: 6
Ні: 2
Потрібні роз'яснення:
0
Ні: 6
Так: 2