МІНІСТЕРСТВО ОСВІТИ І НАУКИ УКРАЇНИ
КАХОВСЬКИЙ ДЕРЖАВНИЙ АГРОТЕХНІЧНИЙ КОЛЕДЖ
Лекція на тему:
«Вегетативні органи рослин. Корінь»
Викладач: Даценко В. В.
Навчально-методична карта заняття № 3.1.
№ спеціальності: 201 «Агрономія»
Навчальна дисципліна: «Ботаніка»
Тема: Корінь.
Мета заняття: Ознайомитись з вегетативними і репродуктивними органами рослин; функціями кореня, його первинною і вторинною будовою.
План та організаційна структура лекції
№ п/п | Елементи заняття; навчальні питання; форми і методи навчання; засоби забезпечення заняття | Доповнення, зміни |
1. | Організаційна частина - перевірка присутності студентів на занятті; - перевірка готовності аудиторії до заняття; - бесіда | 2 хв. |
2. | Актуалізація знань: - бесіда | 2 хв |
3. | Мотивація навчальної діяльності: Корінь відіграє дуже важливу роль і виконує різноманітні функції: поглинає воду, мінеральні і подекуди органічні речовини з грунту і передає їх у стебло, закріплює рослину в субстраті, синтезує деякі органічні речовини, здійснює зв’язок з мікроорганізмами грунту, грибами, бактеріями, є вмістилищем запасних поживних речовин, служить для вегетативного розмноження. - пояснення, бесіда | 2хв. |
4. | Активізація опорних знань:
| 8 хв. |
5. | Повідомлення теми, мети та плану заняття | |
6. | Викладення нового матеріалу: План. 3.1. Корінь 3.1.1. Поняття про вегетативні та генеративні органи. 3.1.2. Морфологічна будова кореня. 3.1.3. Анатомічна будова кореня. 3.1.3.1. Первинна будова. 3.1.3.2. Вторинна будова. 3.1.4. Метаморфози коренів. 3.1.5. Бульбочки. Мікориза.
| 70 хв. |
7. | Закріплення, узагальнення та систематизація навчального матеріалу: - усне фронтальне опитування | 2 хв. |
8. | Підведення підсумків заняття, відповіді на запитання студентів: | 2 хв. |
9. | Домашнє завдання: | 2 хв. |
| В. Г. Хржановський С. Ф. Пономаренко, Ботаніка. К. Вища школа. 1985. с. 53 - 65 | |
Викладач: Даценко В. В.
3. ВЕГЕТАТИВНІ ОРГАНИ РОСЛИН
3.1. Корінь
3.1.1. Поняття про вегетативні та генеративні органи.
3.1.2. Морфологічна будова кореня.
3.1.3. Анатомічна будова кореня.
3.1.3.1. Первинна будова.
3.1.3.2. Вторинна будова.
3.1.4. Метаморфози коренів.
3.1.5. Бульбочки. Мікориза.
3.1.1. Поняття про вегетативні та генеративні органи.
Орган – це відокремлена частина організму, що складається із різних типів клітин і тканин, має стале положення, розвивається у процесі онтогенезу і виконує певну функцію у живому організмі.
В залежності від функцій органи поділяють на вегетативні та генеративні. Вегетативні органи забезпечують життєдіяльність рослини (ріст, розвиток, живлення, обмін речовин, виділення тощо).
До вегетативних органів рослини належить корінь і пагін.
Корінь – це підземна частина рослини. Функції кореня полягають у закріпленні рослини в ґрунті, всмоктуванні води та мінеральних речовин та транспорті їх до надземної частини. Сукупність всіх коренів рослини формує кореневу систему.
Пагін – це надземна частина рослини. Вона складається із осьової частини – стебла, та розміщених на ньому листків і бруньок. (Рис. 3.1.1.1 )
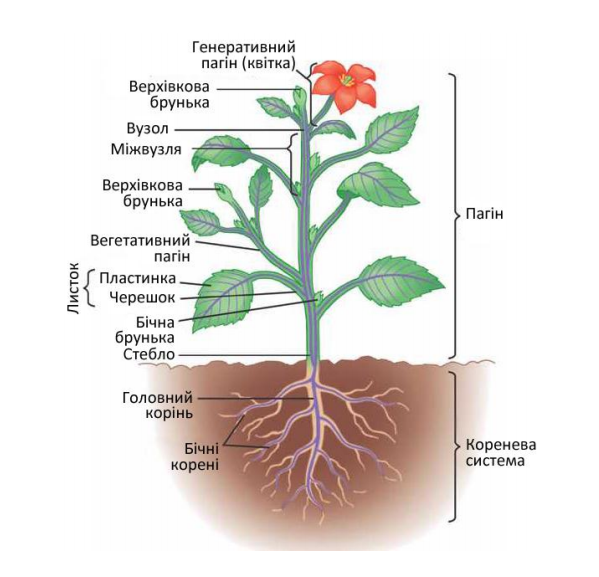 Брунька – це зачаток нового пагона, стебло забезпечує зв’язок між усіма органами рослини, листки здійснюють фотосинтез, газообмін, випаровування води.
Брунька – це зачаток нового пагона, стебло забезпечує зв’язок між усіма органами рослини, листки здійснюють фотосинтез, газообмін, випаровування води.
Для органів рослин характерні деякі закономірності.
Симетрія. Орган називають моносиметричним, якщо через нього можна провести лише одну площину симетрії, полісиметричний, якщо через нього можна провести більш як одну площину симетрії.
Рис.3.1.1.1 Органи рослини.
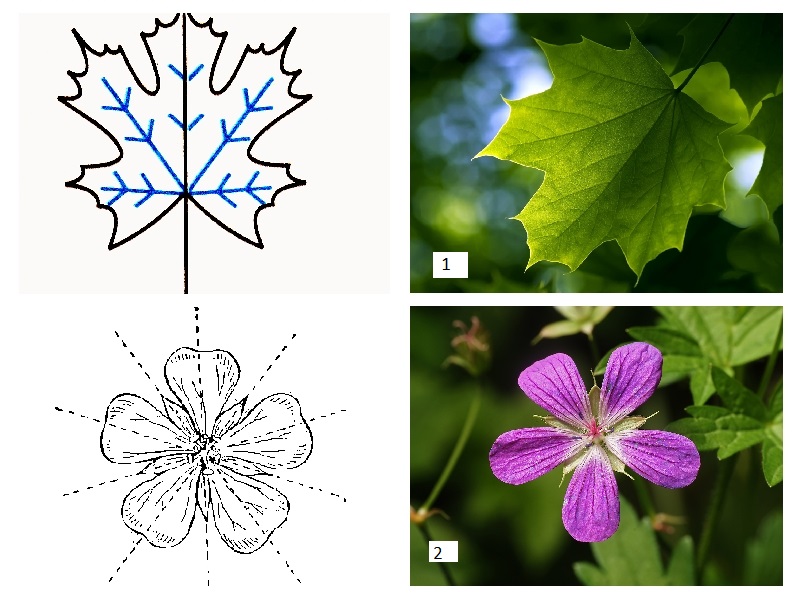
Рис. 3.1.1.2. 1 – моносиметричний орган: 2- полісиметричний орган.
Полярність. Вегетативний орган, або його частина має два полюси: термінальний (верхній) і базальний (нижній). В термінальній частині утворюються лише пагони, а базальній – лише корені.
Геотропізм - здатність органів рости у певному напрямку, спричиненому односторонньою дією сили земного тяжіння. Залежно від напрямку росту органів щодо сили тяжіння виділяють два види геотропізму:
1) позитивний, при якому орган росте до центра землі (за силою земного тяжіння);
2) негативний, при якому орган росте від центру землі (проти сили земного тяжіння).
Особливості розвитку органів односім’ядольних і двосім’ядольних рослин.
Основні вегетативні органи закладені уже у зародку насінини.
За сприятливих умов температури і вологості насіння поглинає воду і, якщо є досить повітря, проростає. Першим проростає корінь, який прориває спермодерму і занурюється у грунт (рис 3.1.1.3.).
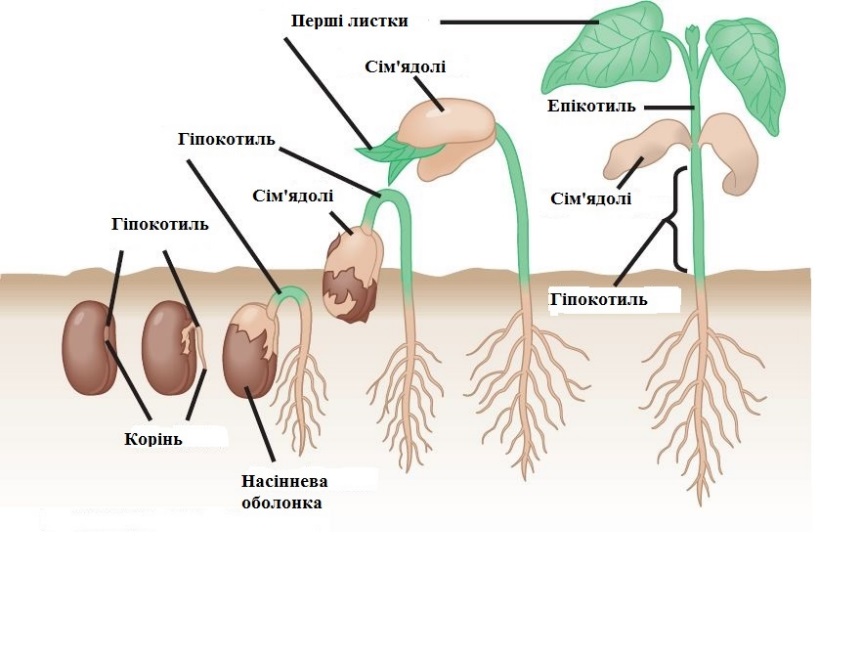
Рис. 3.1.1.3. Проростання насіння квасолі.
Завдяки цьому молодий проросток закріплюється у грунті і поглинає воду з розчиненими в ній солями. Відразу за коренем починає рости пагін. Спочатку він вигнутий і розсуває грунт своїм вигином, потім розправляється і виносить на поверхні сім’ядолі і бруньку. Сім’ядолі зеленіють і певний час виконують функції листків. Брунька продовжує рости вгору, утворюючи стебло і справжні перші листки дорослої рослини. Межу між коренем і стеблом називають кореневою шийкою. Частину стебла між кореневою шийкою і сім’ядолями називають називають - гіпокотелем, а ділянку між сім’ядолями і першим справжнім листком – епікотелем.
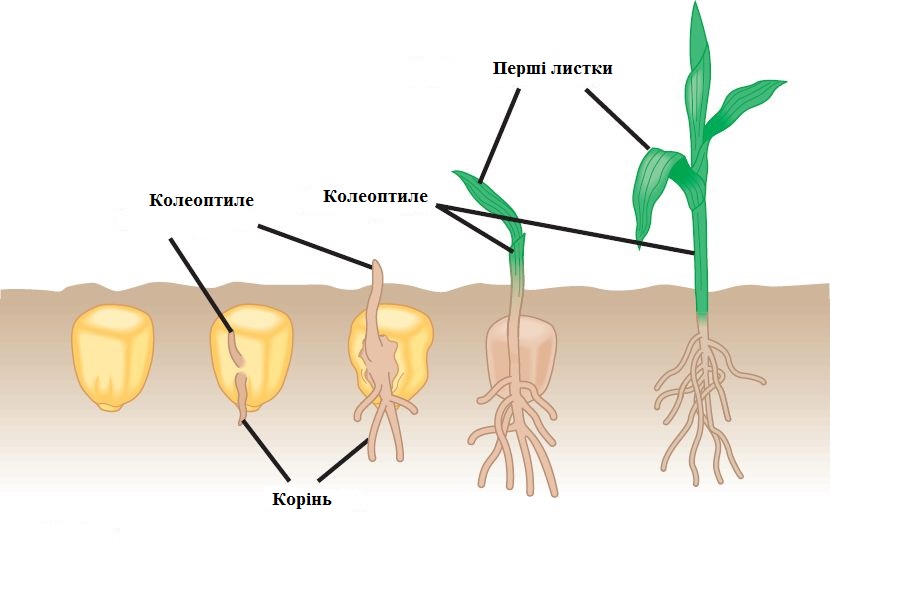
Рис.3.1.1.4. Проростання зернівки пшениці.
3.1.2. Морфологічна будова кореня.
Корінь - підземний осьовий радіально-симетричний вегетативний орган з необмеженим ростом та закріплює рослину в субстраті (ґрунті). Вперше справжні корені з'являються у папоротеподібних. Поява кореня в процесі еволюції рослин - одне із пристосувань до життя на суходолі.
Зона, в якій головний корінь переходить в стебло називається кореневою шийкою. Ріст коренів у рослин триває досить довго. Головний корінь займає вертикальне положення, заглиблюючись в нижні частини ґрунту. По боках головного кореня з’являють бічні корені, які в свою чергу дають початок бічним кореням другого, третього і т. д. порядків.

За походженням розрізняють типи коренів (рис 3.1.2.1):
головний, який утворюється із зародкового корінця насінини;
додаткові, які закладаються на надземній або підземній частині пагона;
бічні, які ендогенно закладаються на головному, додаткових та бічних коренях нижчого порядку.

Рис. 3.1.2.1. 1- головний корінь, 2 – Додаткові корені, 3 – Бічні корені.
Сукупність усіх коренів однієї рослини формує кореневу систему.
Розрізняють такі типи кореневих систем:
стрижнева коренева система, в якої чітко виділяється за розмірами та вертикальним напрямком росту головний корінь, а бічні корені тоненькі (кріп, морква) ;
мичкувата коренева система, в якої головний корінь чітко не відрізняється від численних майже однакових додаткових коренів, може рано відмирати або зовсім не розвиватися (цибуля, жито, пшениця);

Рис. 3.1.2.2. Типи кореневих систем: 1 – стрижнева коренева система, 2 - мичкувата коренева система.
Корені вищих рослин бувають дуже різноманітними за зовнішніми морфологічними ознаками.
Розрізняють такі форми коренів:
шнуроподібна (верблюжа колючка),
ниткоподібна (жито, пшениця),
веретеноподібна (морква, петрушка),
ріпоподібна (ріпа, буряк)
кореневі бульби (пшінка, жоржина) та ін.
Ступінь розвитку кореневої системи залежить від середовища, в якому зростає рослина.
Якщо рослина росте на щільному ґрунті з низьким вмістом кисню та органічних речовин, тоді 90% коренів зосереджені у поверхневому шарі.
На пухких ґрунтах, багатих на органічні сполуки та кисень корені можуть сягати великої глибини (цукровий буряк – 3м, горох – 2м).
3.1.3. Анатомічна будова кореня.
Вздовж кореня можна виділити кілька зон, що відрізняються за будовою і функціями, їх послідовність, починаючи від кінчика така: кореневий чохлик, зона поділу, зона розтягу, всисна зона, провідна зона (рис.3.1.3.1.)
Кореневий чохлик вкриває верхівку кореня. Він складається із кількох шарів живих клітин і запобігає ушкодженню меристеми кореня. Під час просування кореня вглиб ґрунту внаслідок тертя зовнішні клітини чохлика постійно руйнуються, а твірна тканина постійно утворює нові.
Зона поділу – довжиною 2-3мм, складається із клітин твірної тканини (меристеми), ці клітини постійно діляться, підтримуючи власну популяцію, а також дають початок всім іншим клітинам кореня.
Зона розтягування - в ній відбувається видовження клітин, внаслідок чого кінець кореня просувається далі в ґрунт. У верхній частині зони розтягу відбувається диференціація клітин. Довжина цієї ділянки становить кілька міліметрів.
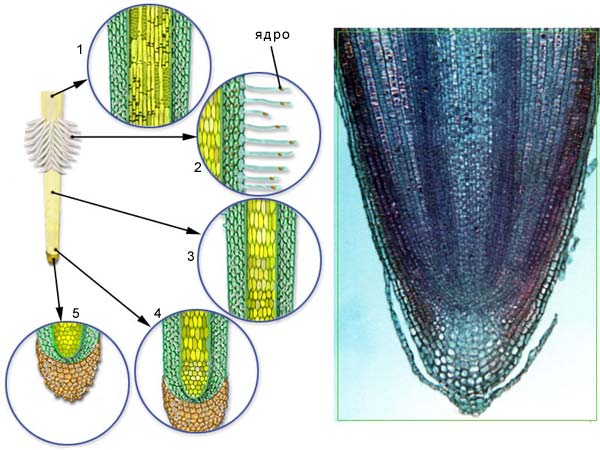
Рис. 3.1.3.1. Зони кореня: 1 – зона проведення, 2 – зона всмоктування, 3 – зона розтягування, 4 – зона поділу клітин, 5 – кореневий чохлик.
Зона всмоктування або зона кореневих волосків розміщена над зоною розтягу. Вона має довжину 5-20мм і щільно вкрита кореневими волосками. На 1мм2 поверхні кореня їх може бути декілька сотень, а на всіх коренях дорослої рослини – кілька мільярдів. Довжина кожного волоска коливається від кількох міліметрів до одного сантиметра. Кореневі волоски живуть недовго – до 20 діб, нові утворюються із верхніх клітин зони розтягу, тому зона всмоктування заглиблюється в ґрунт одночасно із ростом кореня.
Провідна зона або зона бічних коренів. У своїй верхній частині вона межує із основою стебла, яка називається кореневою шийкою.
Основна функція провідної частини кореня – транспорт речовин, всмоктування тут не відбувається. У цій зоні також відбувається галуження кореня.
3.1.3.1. Первинна будова.
Первинна будова кореня - будова, при якій функціонують первинні меристеми. Первинна будова кореня властива однодольним рослинам протягом усього життя та всім голонасінним і дводольним рослинам у молодих ділянках кореня до початку функціонування вторинних меристем (рис.3.1.2.1.1). При первинній будові кореня на поперечному зрізі виділяють такі ділянки:
1. Ризодерма (епіблема) - первинна покривна тканина, клітини якої утворюють кореневі волоски.
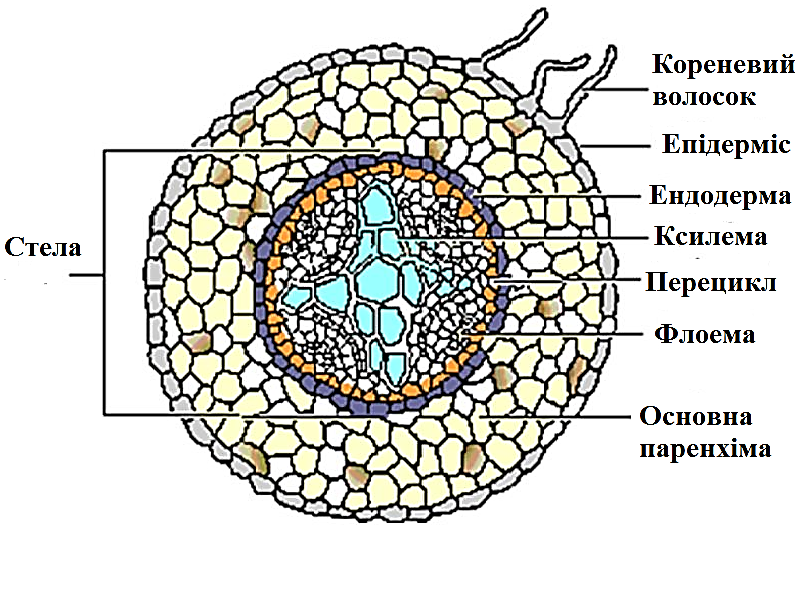
Рис. 3.1.3.1.1. Первинна будова кореня
2. Первинна кора - багатошарова тканина, яка складається з:
а) екзодерми - шар великих багатокутних живих клітин, щільно з' єднаних одна з одною радіальними стінками, які виконують функції захисту і пропускання води і солей.
б) мезодерми - багатошарова паренхіма, яка складається з живих клітин;
в) ендодерми - один шар щільно зімкнених клітин, стінки яких поступово корковіють за рахунок суберину (тому вони не пропускають водє і гази); між цими клітинами розташовані живі тонкостінні пропускні клітини, крізь які проходить вода з розчиненими речовинами з первинної кори до ксилеми центрального циліндра.
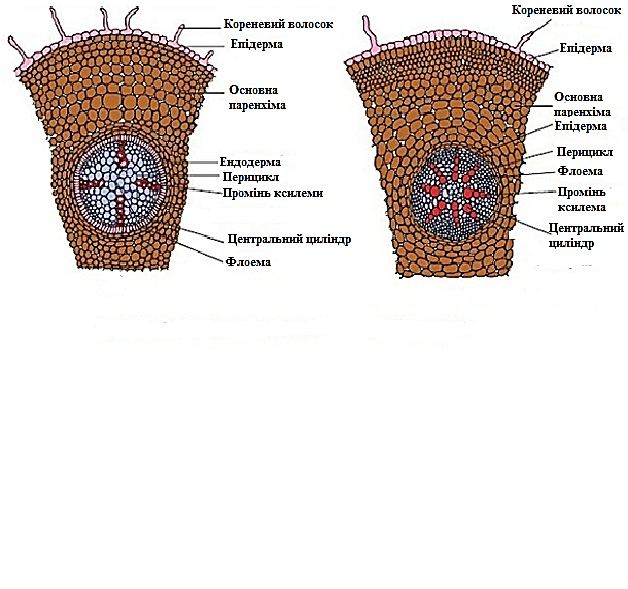
Рис. 3.1.3.1.2. Первинна будова кореня однодольної та двосім’ядольної рослин.
3. Центральний циліндр, або стела складається з різних тканин:
а) перицикл (перикамбій) - шар живих тонкостінних паренхімних клітин, які періодично діляться і дають початок бічним кореням та іншим тканинам (вторинна твірна тканина);
б) паренхімна тканина, в якій радіально розміщений судинний пучок, що складається із ксилеми і флоеми; флоема і ксилема розміщуються на різних радіусах (ксилема ближче до центру).
3.1.3.2. Вторинна будова.
Вторинна будова кореня формується з первинної у результаті діяльності вторинних меристем - камбію і фелогену (рис. 3.1.3.2.1).
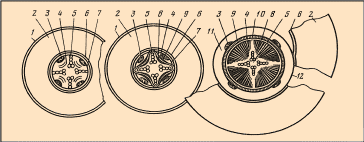
Рис. 3.1.3.2.1. Перехід кореня дводольної рослини від первинної будови до вторинної: 1 – епіблема, 1- первинна кора, 3 – первинна флоема, 4 – камбій, 5 – первинна ксилема, 6- ендодерма, 7 – перицикл, 8 – вторинна ксилема, 9 – вторинна флоема, 10 – радіальний промінь, 11 – паренхіма вторинної кори, 12 – перидерма.
Камбій - вторинна латеральна твірна тканина голонасінних і дводольних рослин. У корені камбій утворюється з паренхімних клітин на межі флоеми і ксилеми, рідше з клітин перициклу. Клітини камбію діляться паралельно поверхні органу, утворюючи з периферійного (зовнішнього) боку елементи вторинної флоеми, а з центрального (внутрішнього) - елементи вторинної ксилеми. Це приводить до потовщення кореня ( рис. 3.1.3.2.2).
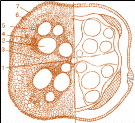
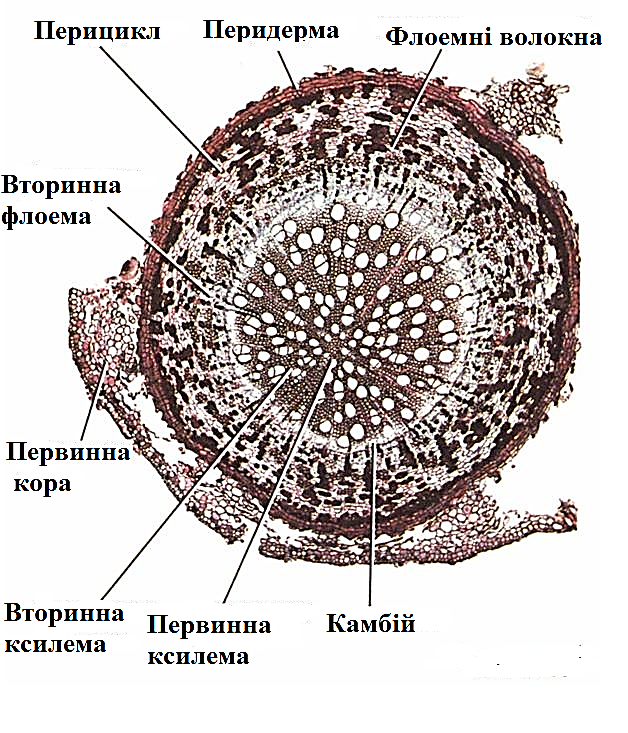
Рис. 3.1.3.2.2. Вторинна будова кореня гарбуза. 1 – первинна ксилема, 2 – вторинна ксилема, 3 – радіальний промінь, 4 – камбій, 5 – первинна і вторинна флоема, 6 – основна паренхіма вторинної кори, 7 - корок.
Поступово із наростанням вторинної ксилеми і вторинної флоеми первинна флоема відтісняється до периферії, а первинна ксилема - до центру.
Фелоген або корковий камбій, - вторинна латеральна меристема, яка закладається або безпосередньо під епідермою, або в глибших шарах первинної кори. Таким чином, на зміну первинній формується вторинна кора, що являє сукупність тканин розташованих зовні від камбію.
Отже, за вторинної будови корінь складається з центрального циліндра, вкритого корком.
3.1.4. Метаморфози коренів.
Крім основних функцій корені можуть виконувати і додаткові, для цього вони набувають особливої будови.
Видозміни кореня бувають підземними і надземними.
До підземних видозмін кореня належать коренеплоди та бульбокорені.
Коренеплід – це потовщений головний корінь рослини, в якому у великій кількості відкладаються поживні речовини. Коренеплоди можна знайти в таких рослин як морква, буряк, редька, петрушка, хрін тощо (рис. 3.1.4.1).

Рис. 3.1.4.1. Коренеплоди: 1 – морква, 2 – редька, 3 – петрушка, 4 – буряк.
Поживні речовини також можуть відкладатись у додаткових або бічних коренях, така видозміна кореня називається бульбокоренем або кореневою бульбою (рис. 3.1.4.2.). До рослин, що утворюють кореневі бульби належать жоржини, пшінка, батат.

Рис. 3.1.4.2. Бульбокорені: 1 – пшінка весняна, 2 – жоржина, 3 – батат, 4 – любка дволиста.
Ходульні корені характерні для представників мангрової рослинності, які ростуть у припливно- відпливній смузі морського берегу (рис. 3.1.4.3).
За походженням ці корені є додатковими, і ростуть з стовбуру та нижніх гілок, в напрямку до землі. Вони слугують рослині для опори та живлення. Подібну функцію виконують додаткові корені кукурудзи. Дуже добре розвинені ходульні корені фікуса - баньяна, їх називають коренями підпірками.

Рис. 3.1.4.3. Ходульні корені: 1- панданус, 2- кукурудза, 3 – мангри, 4 – баньян.
Дихальні корені потрібні рослинам для дихання, тобто забезпечення клітин кореневої системи киснем. Вони утворюються в рослин, що живуть на болотах і надмірно зволожених ґрунтах (рис. 3.1.4.4.).
Д ихальні корені відходять від бічних і піднімаються горизонтально вгору, вони мають тонку кору, через яку може відбуватись газообмін.
ихальні корені відходять від бічних і піднімаються горизонтально вгору, вони мають тонку кору, через яку може відбуватись газообмін.
Рис. 3.1.4.4. Дихальні корені: 1 – авіценія, 2 – кипарис болотяний, 3 - чорні мангри.
Повітряні корені розвиваються у рослин, що ростуть не в ґрунті, а на деревах (рослини епіфіти). Повітряні корені – це додаткові корені, що відходять від стебла і звисають у повітрі (рис. 3.1.4.5).

Рослини-епіфіти із повітряними коренями розповсюджені у тропічних лісах, де в повітрі завжди багато вологи. Зовні повітряні корені вкриті кількома шарами мертвих клітин, в оболонках яких є пори, через які рослина може поглинати воду прямо із повітря.
Рис. 3.1.4.5. Повітряні корені: 1 – орхідея, 2 – бромелія, 3 - катлея
А  симіляційні корені. У багатьох тропічних орхідей епіфітів, які зростають на стовбурах інших дерев подібно до лишайників, утворюються плоскі стрічкоподібні корені. Нижня сторона цих коренів вкрита волосками і поглинає воду з розчиненими солями, а верхня має зелений колір і виконує функції фотосинтезу (рис. 3.1.4.6). Зелені асиміляційні корені має і водяний горіх. Ці корені формуються на підсім'ядольному коліні у вигляді занурених у воду ниток
симіляційні корені. У багатьох тропічних орхідей епіфітів, які зростають на стовбурах інших дерев подібно до лишайників, утворюються плоскі стрічкоподібні корені. Нижня сторона цих коренів вкрита волосками і поглинає воду з розчиненими солями, а верхня має зелений колір і виконує функції фотосинтезу (рис. 3.1.4.6). Зелені асиміляційні корені має і водяний горіх. Ці корені формуються на підсім'ядольному коліні у вигляді занурених у воду ниток
Рис. 3.1.4.6. Асиміляційні корені: 1 – водяний горіх, 2 – орхідеї.
Ч  іпкі корені
іпкі корені
Рослини-ліани, тобто рослини, що мають тонке довге стебло і піднімаються вгору, в'ючись по основі, можуть прикріплюватись до поверхні за допомогою спеціальних додаткових коренів, які називаються чіпкими (рис. 3.1.4.7.).
Рис. 3.1.4.7. Чіпкі корені: 1 – кампсис, 2 – партеноцисус, 3 – плющ.
Корені присоски. Рослини паразити живляться поглинаючи поживні речовини із інших рослин, для цього в них є спеціальні корені-присоски (рис. 3.1.4.8).
Корені-присоски проникають в товщу стебла рослини-живителя і поглинають звідти поживні речовини.
Прикладом рослини-паразита є повитиця. Омела, яку можна побачити на деревах є напівпаразитичною рослиною, оскільки вона, на відміну від повитиці, зберегла здатність до фотосинтезу, про що можна судити із її зеленого забарвлення.

Рис. 3.1.4.8. Корені присоски: 1 – вовчок, 2 – повитиця, 3 - омела.
3.1.5. Бульбочки. Мікориза.
Бульбочки
Бульбочкові бактерії відносяться до роду Rhizobium. Вони мають властивість фіксувати азот із атмосферного повітря і синтезувати органічні азотовмісні сполуки. Ці мікроорганізми утворюють на коренях деяких бобових рослин бульбочки, вступаючи в симбіоз (рис. 3.1.5.1). Дані бактерії переводять азот в сполуки, легко доступні для засвоєння рослинами, а квіткові рослини, в свою чергу, є джерелами живильних речовин для бульбочкових бактерій. Також даний вид бактерій є важливою ланкою в процесі збагачення грунту азотом.

Рис. 3.1.5.1. Бульбочки на коренях бобових.
Бульбочкові бактерії живуть на коренях у 10% рослин із родини бобових. Причому різні види бактерій розвиваються на кореневій системі певних вищих рослин. У віки, кормових бобів, гороху - Rh. Leguminosarum, у буркуну, люцерни - Rhizobium meliloti, у сої - Rh. Japonicum, у конюшини - Rh. Trifolii. Якщо коріння бобових відмирають, а бульби руйнуються, бульбочкові бактерії не гинуть, а ведуть спосіб життя сапрофітів.
Мікориза
Мікориза являє собою симбіоз (взаємовигідне співіснування) міцелію гриба та кореня вищої рослини. У формуванні мікоризи беруть участь з одного боку всі голонасінні рослини і деякі квіткові, а з іншого - такі групи грибів, як базидіоміцети, зигоміцети, аскоміцети, інші (рис. 3.1.5.2 ). Завдяки грибам, збільшується поверхня всмоктування кореневої системи, також з'єднання мінеральних речовин надходять всередину кореня в легко засвоюваній формі . Гриб, у свою чергу, харчується вуглеводами, фітогормонами, амінокислотами, які отримуються з кореня вищої рослини.
Існує три види мікоризи:
ендотрофна,
ектотрофна,
ектоендотрофна.
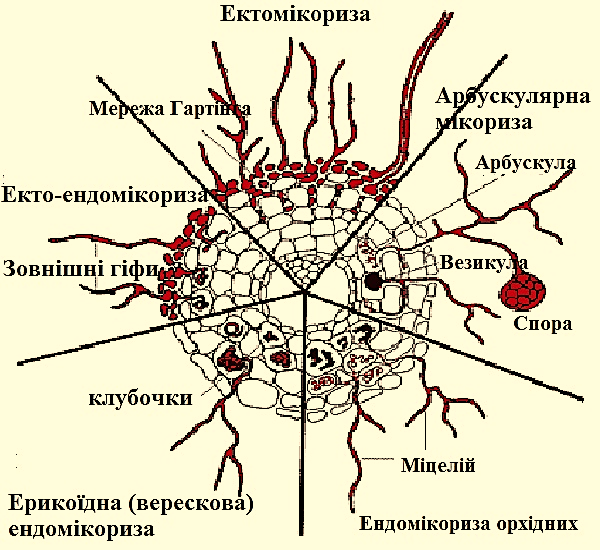
Рис. 3.1.5.2 Види мікоризи
При утворенні ектотрофної (зовнішньої) мікоризи міцелій гриба огортає кінці молодих корінців, формуючи подобу чохлів, і проникає в міжклітинні простори, не руйнуючи клітини. При цьому кореневі волоски відсутні, а кореневий чохлик перетворюється в один-два шари клітин. Корінь виявляється розділеним гіфами гриба на відділи. Таку мережу гіф називають мережею Гартіга. Ектотрофна мікориза спостерігається у багатьох дерев (ялина, дуб, береза), чагарників (верба), зрідка у трав (живородна гречка). Цей тип мікоризи утворюють в більшості випадків гриби гіменоміцети, іноді гастероміцети.
Ендотрофна мікориза характеризується тим, що форма коренів залишається постійною, кореневі волоски зберігаються, немає мережі Гартіга та грибного чохла (рис 3.1.5.3). Гіфи гриба пронизують безпосередньо клітини кореневої паренхіми. Мікориза практично не помітна на поверхні кореня рослини у зв'язку з тим, що значна частина міцелію гриба проникає всередину клітин кореневої системи. У клітинах кореня нагромаджуються грибні гіфи у формі клубків і розгалужених ниток. Мікориза такого типу формується у рослин брусничних, орхідних, , вересових, грушанкових, тощо.

Рис 3.1.5.3 Ендомікориза
При ектоендотрофному типі мікоризи поєднуються властивості екто- та ендомікоріз. Можливо переважання ектотрофного або ендотрофную типу. Така мікориза спостерігається у трав'янистих рослин, чагарників, наприклад, арктоуса арктичного, грушанки крупноквіткової. У цьому випадку гіфи гриба пронизують і клітини кореня, і міжклітинні простори.